Vous aimerez peut-être aussi
- FOTOSSÍNTESEDocument61 pagesFOTOSSÍNTESEClarice GuizoniPas encore d'évaluation
- Extração de Pigmentos FoliaresDocument9 pagesExtração de Pigmentos FoliaresDanielSantosPas encore d'évaluation
- Aula de FotossinteseDocument25 pagesAula de FotossintesePriscila Suelen LopesPas encore d'évaluation
- Metais em Sistemas Aquáticos: Biodisponibilidade e Influência da Matéria OrgânicaD'EverandMetais em Sistemas Aquáticos: Biodisponibilidade e Influência da Matéria OrgânicaPas encore d'évaluation
- Fração Gasosa do Solo: Composição e Importância para o Desenvolvimento das PlantasDocument3 pagesFração Gasosa do Solo: Composição e Importância para o Desenvolvimento das PlantasPatricia MouraPas encore d'évaluation
- Aluno em Foco - Abordando Soluções em Sala de AulaDocument5 pagesAluno em Foco - Abordando Soluções em Sala de Aulavilmar konageski jr100% (1)
- Apostila - ClubedeCinciasGuiadeexperimentoseprticasDocument148 pagesApostila - ClubedeCinciasGuiadeexperimentoseprticasJuliana Schultz100% (1)
- A Temática Ambiental na Escola e os Artefatos da Indústria CulturalD'EverandA Temática Ambiental na Escola e os Artefatos da Indústria CulturalPas encore d'évaluation
- O Mundo Dos Coloides PDFDocument5 pagesO Mundo Dos Coloides PDFMirian AlvesPas encore d'évaluation
- Extração Da ClorofilaDocument8 pagesExtração Da ClorofilaLaise Alves0% (1)
- 004 Aminoacidos Rev PDFDocument2 pages004 Aminoacidos Rev PDFLucas SilvaPas encore d'évaluation
- Experiências de Química Geral by Nito A Debacher, Eduardo StadlerDocument116 pagesExperiências de Química Geral by Nito A Debacher, Eduardo StadlerCristina De Tate SettiPas encore d'évaluation
- FQ 03 C Fisico Quimica SuperiorDocument28 pagesFQ 03 C Fisico Quimica SuperiorLuana GiustoPas encore d'évaluation
- Química II e III - Lista de Exercícios 5 sobre Alcenos e IsomeriaDocument7 pagesQuímica II e III - Lista de Exercícios 5 sobre Alcenos e IsomeriaItalo FerreiraPas encore d'évaluation
- Análises de leite cruDocument13 pagesAnálises de leite cruAmistander SantosPas encore d'évaluation
- O Ensino Das Ciências Através Da Discussão de Controvérsias: Realidade Ou Ficção?Document15 pagesO Ensino Das Ciências Através Da Discussão de Controvérsias: Realidade Ou Ficção?Pedro Rocha dos Reis100% (5)
- Apostila Química AnalitDocument98 pagesApostila Química AnalitFabio SoaresPas encore d'évaluation
- Solução Tampão Ou Sistema TamponanteDocument8 pagesSolução Tampão Ou Sistema TamponanteLeticia SilvaPas encore d'évaluation
- Manual Laboratório Química AnalíticaDocument36 pagesManual Laboratório Química Analítica18taisPas encore d'évaluation
- Exercícios de bioquímica sobre minerais e digestãoDocument18 pagesExercícios de bioquímica sobre minerais e digestãoAline Krei100% (1)
- Análise de Solo para Ciências Agrárias.oDocument174 pagesAnálise de Solo para Ciências Agrárias.ojuniorPas encore d'évaluation
- SOLUÇÃO TAMPÃO - Prática 3Document7 pagesSOLUÇÃO TAMPÃO - Prática 3Leonardo AndradePas encore d'évaluation
- Atividades experimentais para estimular o ensino de Ciências ExatasDocument83 pagesAtividades experimentais para estimular o ensino de Ciências ExatasMarister Oliveira100% (1)
- Determinação de concentração de KMnO4 por espectrofotometriaDocument5 pagesDeterminação de concentração de KMnO4 por espectrofotometriaRansMilerDantasPas encore d'évaluation
- Biologia Celular: Sinalização CelularDocument9 pagesBiologia Celular: Sinalização CelularSandy VanessaPas encore d'évaluation
- Aula HelmintosesDocument69 pagesAula HelmintosesIsabela Costa FurlanPas encore d'évaluation
- ApresentaçãoDocument349 pagesApresentaçãoGabriel100% (1)
- História dos Fogos de ArtifícioDocument51 pagesHistória dos Fogos de ArtifícioGustavo Trindade100% (1)
- Processamento Técnico - HistopatologiaDocument4 pagesProcessamento Técnico - HistopatologiapriscilazevdoPas encore d'évaluation
- Fisiologia Animal ComparadaDocument135 pagesFisiologia Animal ComparadaCaroline Silva Alves100% (2)
- Equilíbrio de PrecipitaçãoDocument29 pagesEquilíbrio de PrecipitaçãoVictor GutierrezPas encore d'évaluation
- Lista de Biologia CelularDocument6 pagesLista de Biologia CelularPriscila OliveiraPas encore d'évaluation
- Análise Química de AlimentosDocument26 pagesAnálise Química de AlimentosLara RangelPas encore d'évaluation
- Membranas CelularDocument29 pagesMembranas CelularLuis Manuel BritoPas encore d'évaluation
- Lista de Exercícios de Matemática (Menos deDocument68 pagesLista de Exercícios de Matemática (Menos deJoão Vitor SoaresPas encore d'évaluation
- Concentração de soluções químicasDocument3 pagesConcentração de soluções químicasEduarda CoelhoPas encore d'évaluation
- Atividade 4 - Saúde - Biologia Celular - 51-2023Document5 pagesAtividade 4 - Saúde - Biologia Celular - 51-2023Cavalini Assessoria AcadêmicaPas encore d'évaluation
- Apostila Ciência e Tecnologia de PolímerosDocument154 pagesApostila Ciência e Tecnologia de PolímerosFernando TertulianoPas encore d'évaluation
- Livro BIO - Química para Ciências BiológicasDocument180 pagesLivro BIO - Química para Ciências Biológicasleonunes80Pas encore d'évaluation
- Estruturas e Componentes Celulares: Uma Visão Microscópica da CélulaDocument57 pagesEstruturas e Componentes Celulares: Uma Visão Microscópica da CélulaBeatriz Fraga0% (1)
- Solubilidade de Compostos OrgânicosDocument7 pagesSolubilidade de Compostos OrgânicosAmanda AguiarPas encore d'évaluation
- História Do MicroscópioDocument4 pagesHistória Do MicroscópioTássia SilveiraPas encore d'évaluation
- Isoterma de Adsorção de Azul de MetilenoDocument23 pagesIsoterma de Adsorção de Azul de MetilenoThamyres CôcoPas encore d'évaluation
- Baixe listas do projeto medicinaDocument13 pagesBaixe listas do projeto medicinaIsadora IsaPas encore d'évaluation
- Diretrizes Curriculares de QuímicaDocument28 pagesDiretrizes Curriculares de QuímicaThais NegrelliPas encore d'évaluation
- Célula eucariótica animal e osmose em hemáciasDocument9 pagesCélula eucariótica animal e osmose em hemáciasEsdras FerrazPas encore d'évaluation
- ProtozoáriosDocument26 pagesProtozoáriosThiago Almeida de Carvalho100% (1)
- Química Geral e InorgânicaDocument20 pagesQuímica Geral e InorgânicaOsmar FagundesPas encore d'évaluation
- Características Gerais Da Célula e Suas EstruturasDocument37 pagesCaracterísticas Gerais Da Célula e Suas EstruturasRobert BrennerPas encore d'évaluation
- Aula Questoes BioquimicaDocument57 pagesAula Questoes BioquimicaIsis ZionPas encore d'évaluation
- Engenharia Bioquímica - Aula 04 - Cinética EnzimáticaDocument51 pagesEngenharia Bioquímica - Aula 04 - Cinética EnzimáticaYuri BarreirosPas encore d'évaluation
- GenéticaDocument20 pagesGenéticaMauro Bittencourt100% (2)
- Metabolismo CelularDocument9 pagesMetabolismo CelularCleyton SantosPas encore d'évaluation
- Compostos OrgânicosDocument36 pagesCompostos OrgânicoschrisPas encore d'évaluation
- 5.1 - Modelo de Resumo Expandido 1Document3 pages5.1 - Modelo de Resumo Expandido 1Julio Cesar EvaristoPas encore d'évaluation
- Extração, Separação, Identificação e Fluorescência de Pigmentos CelularesDocument11 pagesExtração, Separação, Identificação e Fluorescência de Pigmentos CelulareselizangelacarPas encore d'évaluation
- Modelo de ResenhaDocument1 pageModelo de ResenhaelizangelacarPas encore d'évaluation
- Aspectos do Aquecimento Global: Política, Economia e SaúdeDocument1 pageAspectos do Aquecimento Global: Política, Economia e SaúdeelizangelacarPas encore d'évaluation
- Observação de LactobacilosDocument9 pagesObservação de Lactobaciloselizangelacar0% (1)
- LISTA de QUESTO Es Bioquimica Modulo 1Document1 pageLISTA de QUESTO Es Bioquimica Modulo 1elizangelacarPas encore d'évaluation
- Lista de Exercicio 2 ModuloDocument4 pagesLista de Exercicio 2 ModuloelizangelacarPas encore d'évaluation
- 5.1 - Modelo de Resumo Expandido 1Document3 pages5.1 - Modelo de Resumo Expandido 1Julio Cesar EvaristoPas encore d'évaluation
- Genetica Molecular Exercicio AlunosDocument8 pagesGenetica Molecular Exercicio AlunoselizangelacarPas encore d'évaluation
- 6 Observacao de Celulas Eucarioticas Celulas Da Epiderme Da CebolaDocument2 pages6 Observacao de Celulas Eucarioticas Celulas Da Epiderme Da CebolaelizangelacarPas encore d'évaluation
- Superfíciecelular OrganizaçãomolecularefunçãoDocument38 pagesSuperfíciecelular OrganizaçãomolecularefunçãoelizangelacarPas encore d'évaluation
- 4 Uso Do Microscopio Atividade PraticaDocument3 pages4 Uso Do Microscopio Atividade PraticaelizangelacarPas encore d'évaluation
- Zuza - Cariotipo HumanoDocument9 pagesZuza - Cariotipo HumanoLustri CPas encore d'évaluation
- Estudo Dirigido Sobre o citoSOLDocument1 pageEstudo Dirigido Sobre o citoSOLelizangelacarPas encore d'évaluation
- Origem das moléculas das membranasDocument1 pageOrigem das moléculas das membranaselizangelacarPas encore d'évaluation
- Anelas Das Celulas EucarioticasDocument1 pageAnelas Das Celulas EucarioticaselizangelacarPas encore d'évaluation
- Divisao de Trabalho Entre As Celulas DiferenciacaoDocument12 pagesDivisao de Trabalho Entre As Celulas DiferenciacaoelizangelacarPas encore d'évaluation
- 12 Atividade Da Catalase peroXISSOMOSDocument4 pages12 Atividade Da Catalase peroXISSOMOSelizangelacarPas encore d'évaluation
- Identificação de grãos de amido em células de batata usando LugolDocument4 pagesIdentificação de grãos de amido em células de batata usando LugolelizangelacarPas encore d'évaluation
- Panhia Das ProteinasDocument6 pagesPanhia Das ProteinaselizangelacarPas encore d'évaluation
- Movimento de ciclose em cloroplastos de Elodea spDocument4 pagesMovimento de ciclose em cloroplastos de Elodea spelizangelacarPas encore d'évaluation
- PEROXISSOMOSDocument12 pagesPEROXISSOMOSelizangelacarPas encore d'évaluation
- Capitulo 3 Membranas Celulares Permeabilidade Das MembranasDocument18 pagesCapitulo 3 Membranas Celulares Permeabilidade Das MembranaselizangelacarPas encore d'évaluation
- Ponentes Quimicos Da Celula PDFDocument23 pagesPonentes Quimicos Da Celula PDFelizangelacarPas encore d'évaluation
- PlementarDocument3 pagesPlementarelizangelacarPas encore d'évaluation
- Osmose em células vegetais: plasmólise e desplasmóliseDocument4 pagesOsmose em células vegetais: plasmólise e desplasmóliseelizangelacarPas encore d'évaluation
- Osmose em células vegetais: plasmólise e desplasmóliseDocument4 pagesOsmose em células vegetais: plasmólise e desplasmóliseelizangelacarPas encore d'évaluation
- Osmose em células vegetais: plasmólise e desplasmóliseDocument4 pagesOsmose em células vegetais: plasmólise e desplasmóliseelizangelacarPas encore d'évaluation
- ApoptoseDocument12 pagesApoptoseelizangelacarPas encore d'évaluation
- PHDA transição cuidados adultosDocument11 pagesPHDA transição cuidados adultosFilipa FreitasPas encore d'évaluation
- Cronograma Construção de Rampa para Balança RodoviáriaDocument1 pageCronograma Construção de Rampa para Balança RodoviáriaDilson JuniorPas encore d'évaluation
- Formação Coroinhas Jesus de NazaréDocument56 pagesFormação Coroinhas Jesus de NazaréMaria I C Prata80% (5)
- Anatomia TireoideeparatireoideDocument38 pagesAnatomia TireoideeparatireoidethiagomedruasPas encore d'évaluation
- Problema Urbano de Luanda: A Questão Do Lixo Na Capital AngolanaDocument7 pagesProblema Urbano de Luanda: A Questão Do Lixo Na Capital AngolanaSaidi CalalaPas encore d'évaluation
- Os Movimentos Espíritas (Ou A Hidra de Lerna Espírita)Document8 pagesOs Movimentos Espíritas (Ou A Hidra de Lerna Espírita)Antonio BuenoPas encore d'évaluation
- IPAPPI SISTEMA GRISA - Paranormalidade Um Potencial MentalDocument13 pagesIPAPPI SISTEMA GRISA - Paranormalidade Um Potencial MentalSandro Zoleti50% (2)
- Seminovos em Belo Horizonte CarbigDocument1 pageSeminovos em Belo Horizonte CarbigVictor SantosPas encore d'évaluation
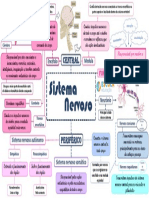
- Sistema nervoso: estrutura e funções principaisDocument1 pageSistema nervoso: estrutura e funções principaisGabriela PiresPas encore d'évaluation
- Curso de Piano - Prefácio e Visão das TeclasDocument1 pageCurso de Piano - Prefácio e Visão das TeclasAmanda PizolPas encore d'évaluation
- FII BM Brascan Lajes Corporativas informativo mensal outubro 2017Document3 pagesFII BM Brascan Lajes Corporativas informativo mensal outubro 2017escobarPas encore d'évaluation
- Macrodantina PDFDocument3 pagesMacrodantina PDFDiego FelixPas encore d'évaluation
- Porifera ResearchDocument694 pagesPorifera ResearchLuiz Mello LulaPas encore d'évaluation
- Geometria analítica: retas e ângulosDocument3 pagesGeometria analítica: retas e ângulosJulio CarraroPas encore d'évaluation
- Relatorio Eng Da Producao e A Profissao (Jonesigley Da Silva Duraes RU 4443925Document2 pagesRelatorio Eng Da Producao e A Profissao (Jonesigley Da Silva Duraes RU 4443925Jonesigley da Silva DurãesPas encore d'évaluation
- Serra Talhada Pereiras e CarvalhoDocument149 pagesSerra Talhada Pereiras e CarvalhoKraus Diniz CoelhoPas encore d'évaluation
- Vim20 Ti P339 07 PortuguesDocument9 pagesVim20 Ti P339 07 PortuguesAndrePas encore d'évaluation
- Manual de Instalação Do Aquecedor SolarDocument6 pagesManual de Instalação Do Aquecedor SolarClaudia Mourão FernandesPas encore d'évaluation
- A Realidade Do Meu Pecado - Salmo 51Document4 pagesA Realidade Do Meu Pecado - Salmo 51gomesdm585100% (1)
- Ajudante de Produção-ServenteDocument1 pageAjudante de Produção-ServenteElisaldo SilvaPas encore d'évaluation
- 2o Encontro Regional História MídiaDocument981 pages2o Encontro Regional História MídiaRodrigoPas encore d'évaluation
- Chave de Nivel Tipo Boia PeraDocument1 pageChave de Nivel Tipo Boia PeraAngelo MachadoPas encore d'évaluation
- CoragemDocument196 pagesCoragemJúlia OhlweilerPas encore d'évaluation
- Rolando Nassau - Ambientes Do Culto IIDocument3 pagesRolando Nassau - Ambientes Do Culto IIIVANTEOLOGIAPas encore d'évaluation
- Análise Linguística 2Document8 pagesAnálise Linguística 2minatinhuhkunPas encore d'évaluation
- Genealogia ConstelacoesDocument50 pagesGenealogia Constelacoesraviresck100% (1)
- As Grandes Profecias Relativas Aos Gentios - G. H. PemberDocument406 pagesAs Grandes Profecias Relativas Aos Gentios - G. H. PemberEDGAR DE PAULO SilvaPas encore d'évaluation
- PRINCÍPIO DA INSIGNIFICÂNCIA X BEM DE VALOR SENTIMENTALDocument3 pagesPRINCÍPIO DA INSIGNIFICÂNCIA X BEM DE VALOR SENTIMENTALCaio MartinsPas encore d'évaluation
- Doenças Crônicas Que Determinam A Realização de Atividades Home OfficeDocument6 pagesDoenças Crônicas Que Determinam A Realização de Atividades Home OfficeAline Sousa JorgePas encore d'évaluation
- Bojana Cvejić - Notas para Uma Sociedade Da Performance - Sobre Dança, Esportes, Museus e Seus UsosDocument17 pagesBojana Cvejić - Notas para Uma Sociedade Da Performance - Sobre Dança, Esportes, Museus e Seus UsosJbrt OuePas encore d'évaluation