Vous aimerez peut-être aussi
- Questions KatzungDocument46 pagesQuestions KatzungYalman Eyinç100% (3)
- Aquaponics DesignDocument74 pagesAquaponics Designwilliam_pepito100% (8)
- Thought Field TherapyDocument5 pagesThought Field TherapyMitz VinegasPas encore d'évaluation
- A Molecular Conception of Organisms and Neoplasms: A Theory That Any Organism Is Basically a Single Chemical MoleculeD'EverandA Molecular Conception of Organisms and Neoplasms: A Theory That Any Organism Is Basically a Single Chemical MoleculePas encore d'évaluation
- English For Physiotherapists 2007 - A.kuciejczykDocument112 pagesEnglish For Physiotherapists 2007 - A.kuciejczykjacekyok2100% (6)
- Apoptosis LectureDocument103 pagesApoptosis LectureGajanan SaykhedkarPas encore d'évaluation
- Best Practice Guide For Environmental Protection On Construction SitesDocument216 pagesBest Practice Guide For Environmental Protection On Construction SitesWL TamPas encore d'évaluation
- Human Chromosomes: An Illustrated Introduction to Human CytogeneticsD'EverandHuman Chromosomes: An Illustrated Introduction to Human CytogeneticsÉvaluation : 5 sur 5 étoiles5/5 (1)
- Sanitary PadDocument5 pagesSanitary PadKenneth GohPas encore d'évaluation
- OkhlaDocument55 pagesOkhlaPoojit Popli50% (2)
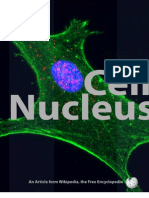
- Cell NucleusDocument17 pagesCell NucleusAsad Aly100% (8)
- Nestle BoycottDocument12 pagesNestle BoycottSubhrajit BhattacharyaPas encore d'évaluation
- Cell StructureandFunctionOfeliaDocument10 pagesCell StructureandFunctionOfeliapiaPas encore d'évaluation
- AnxolamDocument38 pagesAnxolammanjitdeshmukh2Pas encore d'évaluation
- Apoptosis 5Document9 pagesApoptosis 5Kamila Środa-PomianekPas encore d'évaluation
- The Morphology of ApoptosisDocument13 pagesThe Morphology of ApoptosismohammedPas encore d'évaluation
- ApoptosiDocument3 pagesApoptosiMarcoZaccariPas encore d'évaluation
- Historia de La ApoptosisDocument7 pagesHistoria de La ApoptosisAndrea Garcia CaycedoPas encore d'évaluation
- NIH Public Access: Caspase-1 Induced Pyroptotic Cell DeathDocument15 pagesNIH Public Access: Caspase-1 Induced Pyroptotic Cell DeathcutlisaafriannaPas encore d'évaluation
- Original: Yoshiomi TAKAGI, So Kitsunezaki, Tae Ohkido and Rie KOMORIDocument9 pagesOriginal: Yoshiomi TAKAGI, So Kitsunezaki, Tae Ohkido and Rie KOMORIShaker HusienPas encore d'évaluation
- Cell Death and ClearanceDocument41 pagesCell Death and ClearanceZhongming ChenPas encore d'évaluation
- Extrinsic and Intrinsic Apoptosis Signal Pathway Review: Zhao HongmeiDocument20 pagesExtrinsic and Intrinsic Apoptosis Signal Pathway Review: Zhao HongmeiAgustina PramudianingtiasPas encore d'évaluation
- Death Cell JournalDocument13 pagesDeath Cell JournalTirtarizkiPas encore d'évaluation
- J Gerontol A Biol Sci Med Sci-2001-Pollack-B475-82Document8 pagesJ Gerontol A Biol Sci Med Sci-2001-Pollack-B475-82divinebutler13Pas encore d'évaluation
- Bio 2010 60 1 12Document3 pagesBio 2010 60 1 12rmdowney1765Pas encore d'évaluation
- Classification of Cell Death: Fevziye Vural, Suna Cebesoy and Mehmet KarakasDocument7 pagesClassification of Cell Death: Fevziye Vural, Suna Cebesoy and Mehmet KarakasSyahroniPas encore d'évaluation
- Mi 04 005Document11 pagesMi 04 005Jefferson CaesarioPas encore d'évaluation
- The Illusion of Cell ImmortalityDocument6 pagesThe Illusion of Cell ImmortalityMediatheek SGAPas encore d'évaluation
- Exercise # 2: Cells and Cell DivisionDocument10 pagesExercise # 2: Cells and Cell DivisionLiv LeysonPas encore d'évaluation
- Trager 1963Document6 pagesTrager 1963IñigoGaitanSalvatellaPas encore d'évaluation
- Cell Theory, Specificity, and Reproduction, 1837-1870: Europe PMC Funders GroupDocument10 pagesCell Theory, Specificity, and Reproduction, 1837-1870: Europe PMC Funders GroupSilvio SánchezPas encore d'évaluation
- Arcy 2019 Cell Death A Review of The Major Forms of Apoptosis Necrosis and AutophagyDocument11 pagesArcy 2019 Cell Death A Review of The Major Forms of Apoptosis Necrosis and AutophagyCésar HaroPas encore d'évaluation
- Exercise # 2: Cells and Cell DivisionDocument10 pagesExercise # 2: Cells and Cell DivisionEnyong LumanlanPas encore d'évaluation
- Aging Guides: Cellular SenescenceDocument6 pagesAging Guides: Cellular SenescenceKana FajarPas encore d'évaluation
- General Biology Outline 1ST Quarter 1ST SemesterDocument21 pagesGeneral Biology Outline 1ST Quarter 1ST SemesterPrince Caezar BumatayPas encore d'évaluation
- Cell J 13 65 2 PDFDocument8 pagesCell J 13 65 2 PDFRaissaPas encore d'évaluation
- Intreeso To ApoptossisDocument9 pagesIntreeso To ApoptossisToh Qin KanePas encore d'évaluation
- Apoptosis: Apoptosis (From Ancient GreekDocument26 pagesApoptosis: Apoptosis (From Ancient Greekuci kinasePas encore d'évaluation
- Cell Death History and FutureDocument12 pagesCell Death History and FutureFarooqPas encore d'évaluation
- Edited Homeostatis and Death CellDocument34 pagesEdited Homeostatis and Death CellAfridaPas encore d'évaluation
- Programmed Cell Death in AgingDocument25 pagesProgrammed Cell Death in AgingGustavo Sá MottaPas encore d'évaluation
- Programmed Cell Death in Plants: Narcin Palavan-Unsal, Elif-Damla Buyuktuncer and Mehmet Ali TufekciDocument23 pagesProgrammed Cell Death in Plants: Narcin Palavan-Unsal, Elif-Damla Buyuktuncer and Mehmet Ali TufekciSourav GuptaPas encore d'évaluation
- Biology Review Part 3: Cellular Senescence and Medical ImplicationDocument9 pagesBiology Review Part 3: Cellular Senescence and Medical Implicationdjxela89Pas encore d'évaluation
- Atlas of ApoptosisDocument14 pagesAtlas of ApoptosisAnonymous VECoBcRSPas encore d'évaluation
- A Long Twentieth Century of The Cell Cycle and BeyDocument8 pagesA Long Twentieth Century of The Cell Cycle and BeyVidhi SaxenaPas encore d'évaluation
- Cell Senescence & Cell DeathDocument44 pagesCell Senescence & Cell DeathYogesh Bhatt BhattPas encore d'évaluation
- CellDocument3 pagesCellFrama Intan MiguelPas encore d'évaluation
- About The Various Definitions of Life - Pier Luigi LuisiDocument10 pagesAbout The Various Definitions of Life - Pier Luigi LuisiKris Cowell LlorensPas encore d'évaluation
- UPCAT REVIEWER - ScienceDocument4 pagesUPCAT REVIEWER - ScienceNicole de los ReyesPas encore d'évaluation
- ApoptosisDocument1 pageApoptosisMarcus PirotePas encore d'évaluation
- Jilbert B. Gumaru, LPTDocument85 pagesJilbert B. Gumaru, LPTJulius MacaballugPas encore d'évaluation
- Cellular Lifespan and Senescence A Complex BalanceDocument12 pagesCellular Lifespan and Senescence A Complex Balanceprabha karanPas encore d'évaluation
- The Botanical Review 1940 v.06Document63 pagesThe Botanical Review 1940 v.06alborewimi09Pas encore d'évaluation
- Biomol - Lecture 13 - ApoptosisDocument61 pagesBiomol - Lecture 13 - Apoptosismy evePas encore d'évaluation
- Introduction To The Cell and Its StructureDocument6 pagesIntroduction To The Cell and Its StructureMary Ann Leona SelgaPas encore d'évaluation
- PIIS0092867405002333Document3 pagesPIIS0092867405002333Who ?Pas encore d'évaluation
- Cell Theory RevisedDocument24 pagesCell Theory RevisedCésar Octavio Moreno ZúñigaPas encore d'évaluation
- Caveolin-Deficient Mice: Insights Into Caveolar Function Human DiseaseDocument10 pagesCaveolin-Deficient Mice: Insights Into Caveolar Function Human DiseaseStefania CristinaPas encore d'évaluation
- Chapter 1 - Molecular Mechanisms of Cell Death - 2009 - Molecular PathologyDocument22 pagesChapter 1 - Molecular Mechanisms of Cell Death - 2009 - Molecular PathologySELPas encore d'évaluation
- Apoptosis Versus Necrosis: March 2018Document5 pagesApoptosis Versus Necrosis: March 2018Gandhis Apri WidhayantiPas encore d'évaluation
- Apoptosis and DiseaseDocument8 pagesApoptosis and Diseasejeegnesh pPas encore d'évaluation
- Basvanden Boomgaard ThesisDocument9 pagesBasvanden Boomgaard ThesismedtubeburnerPas encore d'évaluation
- Lesson 1 Cell, Cell Theory, and Cell TypesDocument34 pagesLesson 1 Cell, Cell Theory, and Cell TypesAbubakar DucaysanePas encore d'évaluation
- Cell Death in Health and DiseaseDocument11 pagesCell Death in Health and DiseaseSurya DinataPas encore d'évaluation
- General Biology I ReviewerDocument2 pagesGeneral Biology I ReviewerKathleen Kaye Delos AngelesPas encore d'évaluation
- The Mystery of LifeDocument5 pagesThe Mystery of LifeSamuel Venilde SimoquePas encore d'évaluation
- The Cell CycleDocument5 pagesThe Cell CycleCamille Sison-AlmirolPas encore d'évaluation
- The Cell Cycle PDFDocument5 pagesThe Cell Cycle PDFPraise ChinoringaPas encore d'évaluation
- BitumenDocument13 pagesBitumenmicaziv4786Pas encore d'évaluation
- Current State of Alzheimer'S Disease Research and TherapeuticsDocument382 pagesCurrent State of Alzheimer'S Disease Research and TherapeuticsyusufPas encore d'évaluation
- Renato Carpinito Disciplinary Action by Executive Office of Health and Human Services in Boston, MassachusettsDocument25 pagesRenato Carpinito Disciplinary Action by Executive Office of Health and Human Services in Boston, MassachusettsJonathan WeissPas encore d'évaluation
- Guadalquiver v. Sea Power Shipping Enterprise, Inc. GR No. 226200Document2 pagesGuadalquiver v. Sea Power Shipping Enterprise, Inc. GR No. 226200Maynard Gabayno100% (1)
- Omnibus Rules On Leave PDFDocument2 pagesOmnibus Rules On Leave PDFHeatherPas encore d'évaluation
- A Review of Lablab PurpureusDocument16 pagesA Review of Lablab PurpureusAdilah RoddiPas encore d'évaluation
- VPHin IndiaoieproofDocument17 pagesVPHin Indiaoieproofhari prasadPas encore d'évaluation
- Appendix Framework For Rca and Corrective ActionsDocument18 pagesAppendix Framework For Rca and Corrective ActionsMutati TPas encore d'évaluation
- Work CitedDocument5 pagesWork Citedapi-317144934Pas encore d'évaluation
- General Objecti-Wps OfficeDocument3 pagesGeneral Objecti-Wps OfficeSumit YadavPas encore d'évaluation
- Transformers Fire Protection: October 2017Document5 pagesTransformers Fire Protection: October 2017Hedi Ben MohamedPas encore d'évaluation
- Archeology of AcupunctureDocument8 pagesArcheology of AcupuncturecarloPas encore d'évaluation
- 04.4 Sea April 2021 R4Document205 pages04.4 Sea April 2021 R4Pranay PatelPas encore d'évaluation
- Curie Breakthrough by SlidesgoDocument47 pagesCurie Breakthrough by SlidesgoRai YugiPas encore d'évaluation
- Tourism+promotion+and+development +Cynthia+Nokubonga+DubeDocument139 pagesTourism+promotion+and+development +Cynthia+Nokubonga+DubeArvin Delos ReyesPas encore d'évaluation
- 04 2022 Chicha PDRRMODocument6 pages04 2022 Chicha PDRRMOmikko parelPas encore d'évaluation
- Cavite State University: The Fire Safety Measures of Informal Settlers in Barangay Niog 1 Bacoor, Cavite"Document9 pagesCavite State University: The Fire Safety Measures of Informal Settlers in Barangay Niog 1 Bacoor, Cavite"Ralph Joshua BedraPas encore d'évaluation
- Gastroschisis Is A Congenital Anterior Abdominal Wall DefectDocument1 pageGastroschisis Is A Congenital Anterior Abdominal Wall DefectMomogi ForeverhappyPas encore d'évaluation
- Cases Toxo & ForensicDocument27 pagesCases Toxo & ForensicAhmed MohamedPas encore d'évaluation
- EBRSR Handbook Chapter 2 - Brain Reorganization, Recovery and Organized Care - 2020Document64 pagesEBRSR Handbook Chapter 2 - Brain Reorganization, Recovery and Organized Care - 2020anjelikaPas encore d'évaluation
- CNA Body Mechanics Bed Making Comfort CareDocument32 pagesCNA Body Mechanics Bed Making Comfort CareFaith VaughnPas encore d'évaluation