Vous aimerez peut-être aussi
- The Way of The Samurai, Shadowrun BookDocument19 pagesThe Way of The Samurai, Shadowrun BookBraedon Montgomery100% (8)
- Candida Yeast Special Report: Presented by Tony Saldin © 2007 Claire Poulton All Rights ReservedDocument8 pagesCandida Yeast Special Report: Presented by Tony Saldin © 2007 Claire Poulton All Rights ReservedvitallywellPas encore d'évaluation
- HW - 1 SP 2014Document1 pageHW - 1 SP 2014Bhagwan JanePas encore d'évaluation
- Usb Powered Colloidal Silver GeneratorDocument14 pagesUsb Powered Colloidal Silver GeneratorVíctor Faucheret100% (1)
- Chlorine Dioxide Do You Really Need It This Will Help You Decide!Document8 pagesChlorine Dioxide Do You Really Need It This Will Help You Decide!Pristine WaterPas encore d'évaluation
- LDN Letter To Give To Your DoctorDocument3 pagesLDN Letter To Give To Your DoctorbktangoPas encore d'évaluation
- SilverMAX Colloidal Silver GeneratorDocument3 pagesSilverMAX Colloidal Silver GeneratorLiviu AngelescuPas encore d'évaluation
- Wild Oregano BrochureDocument2 pagesWild Oregano Brochurerosnie22Pas encore d'évaluation
- Can Coconuts Save The World? Monolaurin Versus COVID-19Document26 pagesCan Coconuts Save The World? Monolaurin Versus COVID-19John RitchiePas encore d'évaluation
- Benefits, Dosage, and Side EffectsDocument4 pagesBenefits, Dosage, and Side EffectspartaiePas encore d'évaluation
- Mono LaurinDocument24 pagesMono LaurinMey Shelly RikinPas encore d'évaluation
- Colloidal Silver and Its Amazing Health Benefits (Johan Lööf)Document67 pagesColloidal Silver and Its Amazing Health Benefits (Johan Lööf)Jac100% (1)
- 2014-07-20 George Lea Web99 - Routines and Prep..Document41 pages2014-07-20 George Lea Web99 - Routines and Prep..Lisa KrissPas encore d'évaluation
- Diatomaceous EarthDocument2 pagesDiatomaceous EarthnicosiacyprusPas encore d'évaluation
- Zapper MejoradoDocument2 pagesZapper MejoradoicewarPas encore d'évaluation
- Natural ProductsDocument8 pagesNatural ProductsPraveen T MPas encore d'évaluation
- Cranial Electrotherapy Stimulation: Its First Fifty Years, Plus ThreeDocument36 pagesCranial Electrotherapy Stimulation: Its First Fifty Years, Plus ThreeSilambarasu KaruppiahPas encore d'évaluation
- Beck Protocol Newsletter Nov 02Document4 pagesBeck Protocol Newsletter Nov 02inalight100% (1)
- Clo2 DIY Spreadsheets (MMS - CDS)Document1 pageClo2 DIY Spreadsheets (MMS - CDS)bagus918Pas encore d'évaluation
- Coconut Oil Offers Hope For Antibiotic Resistant GermsDocument5 pagesCoconut Oil Offers Hope For Antibiotic Resistant GermsLiza Y. EspinosaPas encore d'évaluation
- Mild Silver ProteinDocument1 pageMild Silver ProteinJohn MoylanPas encore d'évaluation
- Hydrogen Peroxide DripDocument13 pagesHydrogen Peroxide DripAya100% (1)
- Introduction To Corrective Health MedicineDocument107 pagesIntroduction To Corrective Health MedicineBradford S. Weeks100% (2)
- What Leading Doctors, Experts & Institutes Say About (Colloidal) SilverDocument58 pagesWhat Leading Doctors, Experts & Institutes Say About (Colloidal) SilverZac IngPas encore d'évaluation
- 2 - What To Use Colloidal Silver For-2016Document24 pages2 - What To Use Colloidal Silver For-2016AB236100% (1)
- HomocysteinDocument2 pagesHomocysteinTunas HarapanPas encore d'évaluation
- The Ultimate Simple Protocol For Stage IV VCancerDocument13 pagesThe Ultimate Simple Protocol For Stage IV VCancerMurphy9190Pas encore d'évaluation
- MMS2 PDFDocument24 pagesMMS2 PDFTejesh PatelPas encore d'évaluation
- Colloidal Silver GuideDocument17 pagesColloidal Silver GuideKaren PolzinPas encore d'évaluation
- General Introduction To AnolyteDocument31 pagesGeneral Introduction To AnolyteAjay SharmaPas encore d'évaluation
- Natures.n1 Healing - SecretDocument29 pagesNatures.n1 Healing - Secretwikorv8497Pas encore d'évaluation
- Dr. Nudrat Fatima: Course Title: PHARMACOGNOSY-IB (Basic) (Theory) Course Code: PHARM 418 Cr. Hr. 03Document22 pagesDr. Nudrat Fatima: Course Title: PHARMACOGNOSY-IB (Basic) (Theory) Course Code: PHARM 418 Cr. Hr. 03bush kamPas encore d'évaluation
- Cancer Cure Bicarb Soda Maple Syrup TonicDocument1 pageCancer Cure Bicarb Soda Maple Syrup Toniclihu74Pas encore d'évaluation
- Weston Price Sobre K2Document104 pagesWeston Price Sobre K2Antonella MarreirosPas encore d'évaluation
- Heavy Metal DetoxDocument17 pagesHeavy Metal Detoxnazziwa mercyPas encore d'évaluation
- CandidaDocument1 pageCandidalopansPas encore d'évaluation
- ToddOvokaitys PDFDocument12 pagesToddOvokaitys PDF928371100% (1)
- Cancer Cure Booklet - Drkelleyenzymes PDFDocument21 pagesCancer Cure Booklet - Drkelleyenzymes PDFSengPas encore d'évaluation
- Nature's Cancer Prevention - Vitamin B17Document3 pagesNature's Cancer Prevention - Vitamin B17cohenfuPas encore d'évaluation
- Colloidal Silver Cures MRSA ReportDocument16 pagesColloidal Silver Cures MRSA ReportGodwinPas encore d'évaluation
- Cold and Flu ReliefDocument2 pagesCold and Flu ReliefTheVitaminStore.com100% (1)
- Can Bacteria Cause CancerDocument6 pagesCan Bacteria Cause CancerRon FalconePas encore d'évaluation
- IC FinalDocument64 pagesIC FinalRichfelPas encore d'évaluation
- CDH4000 Combined Formulation, Dilution & Use Instr. Ver 1.4 Including COVID Addition From Dr. Callisperis (Personalized)Document4 pagesCDH4000 Combined Formulation, Dilution & Use Instr. Ver 1.4 Including COVID Addition From Dr. Callisperis (Personalized)Scott McRaePas encore d'évaluation
- CDS-how To Make 3000 PPM Using 4pc HCLDocument2 pagesCDS-how To Make 3000 PPM Using 4pc HCLbagus918Pas encore d'évaluation
- Iodine Summaryupdate 2016Document26 pagesIodine Summaryupdate 2016FrankPas encore d'évaluation
- Use of Coconut OilDocument5 pagesUse of Coconut OilRakeshKumarPas encore d'évaluation
- Beta Glucan InteractionsDocument2 pagesBeta Glucan InteractionsSASA100% (1)
- Colloidal SilverDocument4 pagesColloidal SilverVince MiltonPas encore d'évaluation
- CalcificationDocument7 pagesCalcificationAdrian Stan100% (1)
- Case Study - High Dose Intravenous Vitamin C in The Treatment of A Patient With Adrenocarcinoma of The KidneyDocument3 pagesCase Study - High Dose Intravenous Vitamin C in The Treatment of A Patient With Adrenocarcinoma of The KidneyFilipos ConstantinPas encore d'évaluation
- What Is CDS?Document1 pageWhat Is CDS?JacksonPas encore d'évaluation
- TD DMPS 1Document8 pagesTD DMPS 1api-3714923Pas encore d'évaluation
- Colloidal SilverDocument25 pagesColloidal SilverabazanPas encore d'évaluation
- Hooper David - The Colloidal Silver Hand BookDocument23 pagesHooper David - The Colloidal Silver Hand BookStarla ShermanPas encore d'évaluation
- Cholesterol Cure: Heal Naturally, Without MedicationD'EverandCholesterol Cure: Heal Naturally, Without MedicationPas encore d'évaluation
- Colloidal Silver - The Natural Antibiotic: The Holistic Alternative To Antibiotics New DiscoveredD'EverandColloidal Silver - The Natural Antibiotic: The Holistic Alternative To Antibiotics New DiscoveredPas encore d'évaluation
- RRC & RabDocument14 pagesRRC & RabSyed Waqas AhmedPas encore d'évaluation
- User Manual - Wellwash ACDocument99 pagesUser Manual - Wellwash ACAlexandrPas encore d'évaluation
- Omegas Prezentacija 01Document20 pagesOmegas Prezentacija 01Predrag Djordjevic100% (1)
- Applications of Remote Sensing and Gis For UrbanDocument47 pagesApplications of Remote Sensing and Gis For UrbanKashan Ali KhanPas encore d'évaluation
- What Is A Timer?Document12 pagesWhat Is A Timer?Hemraj Singh Rautela100% (1)
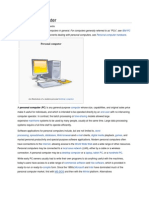
- Personal ComputerDocument3 pagesPersonal ComputerDan Mark IsidroPas encore d'évaluation
- Flyweis Services Pvt. LTDDocument11 pagesFlyweis Services Pvt. LTDFlyweis TechnologyPas encore d'évaluation
- Toeic VocabularyDocument10 pagesToeic VocabularyBrian Niblo80% (5)
- The Accreditation Committee Cityland Development CorporationDocument5 pagesThe Accreditation Committee Cityland Development Corporationthe apprenticePas encore d'évaluation
- Problem SetsDocument69 pagesProblem SetsAnnagrazia ArgentieriPas encore d'évaluation
- Reverse LogisticsDocument37 pagesReverse Logisticsblogdogunleashed100% (7)
- GRADE 1 MUSIC Week 1 Learning PacketsDocument16 pagesGRADE 1 MUSIC Week 1 Learning PacketsQuennie Rose EderPas encore d'évaluation
- CCBA Exam: Questions & Answers (Demo Version - Limited Content)Document11 pagesCCBA Exam: Questions & Answers (Demo Version - Limited Content)begisep202Pas encore d'évaluation
- MSC 200Document18 pagesMSC 200Amit KumarPas encore d'évaluation
- Aruba 8325 Switch SeriesDocument51 pagesAruba 8325 Switch SeriesgmtrlzPas encore d'évaluation
- Filtomat M300Document4 pagesFiltomat M300Sasa Jadrovski100% (1)
- Indiana Property Tax Benefits: (This Form Must Be Printed On Gold or Yellow Paper)Document2 pagesIndiana Property Tax Benefits: (This Form Must Be Printed On Gold or Yellow Paper)abramsdcPas encore d'évaluation
- UTM Firewalls: AR3050S and AR4050SDocument8 pagesUTM Firewalls: AR3050S and AR4050SChristian Javier Lopez DuranPas encore d'évaluation
- E-Cat35xt014 Xtro PhantomsDocument32 pagesE-Cat35xt014 Xtro PhantomsKari Wilfong100% (5)
- ECDIS Presentation Library 4Document16 pagesECDIS Presentation Library 4Orlando QuevedoPas encore d'évaluation
- ALA - Assignment 3 2Document2 pagesALA - Assignment 3 2Ravi VedicPas encore d'évaluation
- Local, Local Toll and Long Distance CallingDocument2 pagesLocal, Local Toll and Long Distance CallingRobert K Medina-LoughmanPas encore d'évaluation
- FoodhallDocument3 pagesFoodhallswopnilrohatgiPas encore d'évaluation
- Cover Sheet: Online Learning and Teaching (OLT) Conference 2006, Pages Pp. 21-30Document12 pagesCover Sheet: Online Learning and Teaching (OLT) Conference 2006, Pages Pp. 21-30Shri Avinash NarendhranPas encore d'évaluation
- GlobalDocument24 pagesGloballaleye_olumidePas encore d'évaluation
- Ansi Numerical CodeDocument6 pagesAnsi Numerical Codekachra13Pas encore d'évaluation
- Form IEPF 2 - 2012 2013FDocument2 350 pagesForm IEPF 2 - 2012 2013FYam ServínPas encore d'évaluation
- Chapter-4 Conditional and Iterative Statements in PythonDocument30 pagesChapter-4 Conditional and Iterative Statements in Pythonashishiet100% (1)
- IFE Level 4 Certificate in Fire InvestigationDocument16 pagesIFE Level 4 Certificate in Fire InvestigationMarlon FordePas encore d'évaluation