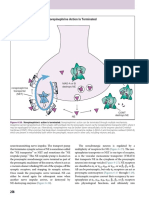
Vous aimerez peut-être aussi
- The Effects of Chronic Corticosterone On PDFDocument5 pagesThe Effects of Chronic Corticosterone On PDFJean Pierre Chastre LuzaPas encore d'évaluation
- Nicotine Potentiates The Antidepressant Effects of Imipramine and Sertraline Sahar M. KamalDocument5 pagesNicotine Potentiates The Antidepressant Effects of Imipramine and Sertraline Sahar M. KamalAlexPas encore d'évaluation
- Ref For Diazepam at ReterivalDocument9 pagesRef For Diazepam at ReterivalKapil SoniPas encore d'évaluation
- TMP - 16938-02JNS - Popoli A2A NP QA-774414356Document9 pagesTMP - 16938-02JNS - Popoli A2A NP QA-774414356Lulu LuwiiPas encore d'évaluation
- 38 Coban Originial 9 3Document7 pages38 Coban Originial 9 3manu choudharyPas encore d'évaluation
- Text S2 Direct Sequencing: Genomic DNA Was Prepared From Collected Blood Samples Using TheDocument8 pagesText S2 Direct Sequencing: Genomic DNA Was Prepared From Collected Blood Samples Using TheRon DSilvaPas encore d'évaluation
- Out 46Document6 pagesOut 46natanatalisPas encore d'évaluation
- Long-Term Potentiation of Intrinsic Excitability at The Mossy Fiber - Granule Cell Synapse of Rat CerebellumDocument9 pagesLong-Term Potentiation of Intrinsic Excitability at The Mossy Fiber - Granule Cell Synapse of Rat CerebellumJar JarPas encore d'évaluation
- Acquisition of Eyeblink Conditioning Is Critically Dependent On Normal Function in Cerebellar Cortical Lobule HVIDocument8 pagesAcquisition of Eyeblink Conditioning Is Critically Dependent On Normal Function in Cerebellar Cortical Lobule HVIOlga Peña HernandezPas encore d'évaluation
- Influence of Different Anaesthetics On Extracellular Aminoacids in Rat BrainDocument5 pagesInfluence of Different Anaesthetics On Extracellular Aminoacids in Rat BrainPau Valle LunaPas encore d'évaluation
- Destructive Pathological Changes in The Rat Spinal Cord Due To Chronic Mechanical CompressionDocument7 pagesDestructive Pathological Changes in The Rat Spinal Cord Due To Chronic Mechanical CompressionasdzxcPas encore d'évaluation
- Effects of Long-Term Acetyl-L-carnitine Administration in Rats: I. Increased Dopamine Output in Mesocorticolimbic Areas and Protection Toward Acute Stress ExposureDocument11 pagesEffects of Long-Term Acetyl-L-carnitine Administration in Rats: I. Increased Dopamine Output in Mesocorticolimbic Areas and Protection Toward Acute Stress ExposureTormentyPas encore d'évaluation
- The Process of Kindling and The Study of Epileptogenic Phenomena Within The Limbic System The Kindling ProcessDocument4 pagesThe Process of Kindling and The Study of Epileptogenic Phenomena Within The Limbic System The Kindling ProcessOwen BunaganPas encore d'évaluation
- Greg D. Gale Et Al - Cholinergic Modulation of Pavlovian Fear Conditioning: Effects of Intrahippocampal Scopolamine InfusionDocument6 pagesGreg D. Gale Et Al - Cholinergic Modulation of Pavlovian Fear Conditioning: Effects of Intrahippocampal Scopolamine InfusionHumiopPas encore d'évaluation
- Cognition-Enhancing and Anxiolytic Effects of Memantine: Rimante Minkeviciene, Pradeep Banerjee, Heikki TanilaDocument7 pagesCognition-Enhancing and Anxiolytic Effects of Memantine: Rimante Minkeviciene, Pradeep Banerjee, Heikki TanilaDian GbligPas encore d'évaluation
- Franscescon Et Al - 2020Document8 pagesFranscescon Et Al - 2020Cecilia DutraPas encore d'évaluation
- AM404 Anandamide Reuptake Inhibition-2pDocument9 pagesAM404 Anandamide Reuptake Inhibition-2pChris OPas encore d'évaluation
- Scirobotics - Abk2378 SMDocument16 pagesScirobotics - Abk2378 SMxiaohangchiPas encore d'évaluation
- Bloqueo Plexo PDFDocument12 pagesBloqueo Plexo PDFAgustin ColettiPas encore d'évaluation
- 6-Hydroxydopamine Lesion of The Rat Prefrontal CortexDocument5 pages6-Hydroxydopamine Lesion of The Rat Prefrontal CortexJonathanPas encore d'évaluation
- 5990 FullDocument10 pages5990 FullRomina MendezBrandanPas encore d'évaluation
- in (1) Article25Document8 pagesin (1) Article25Ashes To BurnsPas encore d'évaluation
- Ijbms 19 800Document4 pagesIjbms 19 800wineniPas encore d'évaluation
- Ket 3xtDocument9 pagesKet 3xtMiXit LivesPas encore d'évaluation
- Acupuncture Stimulates The Release of Serotonin, But Not Dopamine, in The Rat Nucleus AccumbensDocument6 pagesAcupuncture Stimulates The Release of Serotonin, But Not Dopamine, in The Rat Nucleus Accumbensmarcus_irishPas encore d'évaluation
- Neuropharmacological Activity of Solanum Nigrum FruitDocument6 pagesNeuropharmacological Activity of Solanum Nigrum Fruittanvir.radiant officerPas encore d'évaluation
- ESTRESDocument7 pagesESTRESLUIS FERNANDO DÁVALOS CASTROPas encore d'évaluation
- 1995 Pallarés Nadal Ketamine Lever PessDocument4 pages1995 Pallarés Nadal Ketamine Lever PessRocío Donaire CortésPas encore d'évaluation
- The Authors (2007) - Journal Compilation Federation of European Neuroscience Societies and Blackwell Publishing LTDDocument17 pagesThe Authors (2007) - Journal Compilation Federation of European Neuroscience Societies and Blackwell Publishing LTDAvril NuchePas encore d'évaluation
- Fuentealba J Et Al 2007Document4 pagesFuentealba J Et Al 2007Claudia Perez ManriquezPas encore d'évaluation
- Neutrophil Accumulation After Traumatic Brain Injury in RatsDocument8 pagesNeutrophil Accumulation After Traumatic Brain Injury in RatsNurul WijayantiPas encore d'évaluation
- Pharmacology, Biochemistry and Behavior - Seizure Onset Times For Rats Receiving Systemic Lithium and Pi - 2002 - Persinger, Stewart Et AlDocument11 pagesPharmacology, Biochemistry and Behavior - Seizure Onset Times For Rats Receiving Systemic Lithium and Pi - 2002 - Persinger, Stewart Et AlhimkeradityaPas encore d'évaluation
- GABAp N CognitionDocument5 pagesGABAp N CognitionAsher2225Pas encore d'évaluation
- The Effect of Repeated Exposure To Ethanol On Pre-Existing Fear Memories in RatsDocument8 pagesThe Effect of Repeated Exposure To Ethanol On Pre-Existing Fear Memories in RatsAmanda MageskiPas encore d'évaluation
- Inhibition of NR2B Phosphorylation Restores Alterations in NMDA Receptor Expression and Improves Functional Recovery Following Traumatic Brain Injury in Mice (2008)Document13 pagesInhibition of NR2B Phosphorylation Restores Alterations in NMDA Receptor Expression and Improves Functional Recovery Following Traumatic Brain Injury in Mice (2008)Apróné Török IbolyaPas encore d'évaluation
- Glenn Legault Et Al - Scopolamine During The Paradoxical Sleep Window Impairs Radial Arm Maze Learning in RatsDocument7 pagesGlenn Legault Et Al - Scopolamine During The Paradoxical Sleep Window Impairs Radial Arm Maze Learning in RatsHumiopPas encore d'évaluation
- Does N-Acetylcysteine Have An Effect On Acetylcholine-Induced Contractions and Histopathological Changes On Isolated Rat Ileum?Document5 pagesDoes N-Acetylcysteine Have An Effect On Acetylcholine-Induced Contractions and Histopathological Changes On Isolated Rat Ileum?yannecaPas encore d'évaluation
- NootropicDocument31 pagesNootropicVedanth Reddy100% (2)
- Gumrukcu2017 PDFDocument3 pagesGumrukcu2017 PDFSardjitoPas encore d'évaluation
- Prevention of Laryngospasm With Rocuronium in Cats, A Dose-Finding StudyDocument8 pagesPrevention of Laryngospasm With Rocuronium in Cats, A Dose-Finding StudybrasilvilermandoPas encore d'évaluation
- This Document Contains Text Automatically Extracted From A PDF or Image File. Formatting May Have Been Lost and Not All Text May Have Been RecognizedDocument41 pagesThis Document Contains Text Automatically Extracted From A PDF or Image File. Formatting May Have Been Lost and Not All Text May Have Been RecognizedМатиас Себальос ГусманPas encore d'évaluation
- Tudury, Figueiredo Et AlDocument5 pagesTudury, Figueiredo Et AlZullyPas encore d'évaluation
- Evaluation of Antiepileptic DrugsDocument32 pagesEvaluation of Antiepileptic DrugsRajpal SinghPas encore d'évaluation
- 49 Manuscript 498 1 10 20180701Document5 pages49 Manuscript 498 1 10 20180701Liliana QueiroloPas encore d'évaluation
- OK - Schroeder PDFDocument9 pagesOK - Schroeder PDFFelipe Bueno PatricioPas encore d'évaluation
- Effect of Mast Cell Stabilizer KetotifenDocument9 pagesEffect of Mast Cell Stabilizer KetotifenEvelinaPas encore d'évaluation
- Galati2000 Effects of Teucrium Di6aricatum Heldr. Ssp. Di6aricatumDocument6 pagesGalati2000 Effects of Teucrium Di6aricatum Heldr. Ssp. Di6aricatumDjamila HEZILPas encore d'évaluation
- 5.jan 2017 Melatonin MVPDocument3 pages5.jan 2017 Melatonin MVPTushar BahetiPas encore d'évaluation
- Stroop Macdonald 2000Document4 pagesStroop Macdonald 2000Mar Ruiz CuadraPas encore d'évaluation
- Toggle JurnalDocument2 pagesToggle JurnalAnasta MePas encore d'évaluation
- Growth Hormone Prevents The Memory Deficit Caused by Oxidative Stress in Early Neurodegenerative Stage in RatsDocument7 pagesGrowth Hormone Prevents The Memory Deficit Caused by Oxidative Stress in Early Neurodegenerative Stage in Ratsmada oktav cakradwipaPas encore d'évaluation
- Science - Abi7852 SMDocument41 pagesScience - Abi7852 SMJacob NewellPas encore d'évaluation
- 0717 9502 Ijmorphol 41 04 1184Document7 pages0717 9502 Ijmorphol 41 04 1184paulaalessiaPas encore d'évaluation
- Interim Report Fareks Leonardo DemasDocument3 pagesInterim Report Fareks Leonardo Demas08Leonardo Demas Krisna Satria WijayaPas encore d'évaluation
- Role of Agmatine On Impaired Hippocampal Memory Processing Insulin Resistant Diabetic RatsDocument11 pagesRole of Agmatine On Impaired Hippocampal Memory Processing Insulin Resistant Diabetic RatsnikuPas encore d'évaluation
- The Effects of Atropine and Neostigmine On Heart Rate and RhythmDocument9 pagesThe Effects of Atropine and Neostigmine On Heart Rate and RhythmgugicevdzocePas encore d'évaluation
- Acupuncture-Stimulated Activation of Sensory NeuronsDocument8 pagesAcupuncture-Stimulated Activation of Sensory NeuronsSam WardPas encore d'évaluation
- Vutskits 2007Document11 pagesVutskits 2007myobPas encore d'évaluation
- Sodium Butyrate DoseDocument8 pagesSodium Butyrate Doseuppada.89Pas encore d'évaluation
- Individual Differences in Diving - Intelligence, Personality and Underwater Adaptation - Colodro-Plaza2015Document13 pagesIndividual Differences in Diving - Intelligence, Personality and Underwater Adaptation - Colodro-Plaza2015Laura Elena Amaya DuránPas encore d'évaluation
- Nelson 2012Document19 pagesNelson 2012Laura Elena Amaya DuránPas encore d'évaluation
- Estres y Consolidacion C Sandi PDFDocument6 pagesEstres y Consolidacion C Sandi PDFLaura Elena Amaya DuránPas encore d'évaluation
- Atención y Funcionamiento Ejecutivo en La Rehab Neuropsic de Los Procesos VisuoespacialesDocument9 pagesAtención y Funcionamiento Ejecutivo en La Rehab Neuropsic de Los Procesos VisuoespacialesLaura Elena Amaya DuránPas encore d'évaluation
- A Longitudinal Study of Neurological Soft Signs From Late Childhood Into Early AdulthoodDocument6 pagesA Longitudinal Study of Neurological Soft Signs From Late Childhood Into Early AdulthoodLaura Elena Amaya DuránPas encore d'évaluation
- Pharmacology Midterm BlueDocument5 pagesPharmacology Midterm BlueRhealyn LegaspiPas encore d'évaluation
- Autonomic Nervous Syste and Neurohumoral Transmission-Dr - Jibachha Sah, M.V.SC, LecturerDocument32 pagesAutonomic Nervous Syste and Neurohumoral Transmission-Dr - Jibachha Sah, M.V.SC, Lecturerjibachha sahPas encore d'évaluation
- Stahl' Essential Psychopharmacology 276 PDFDocument1 pageStahl' Essential Psychopharmacology 276 PDFMuhammad AzkaPas encore d'évaluation
- Katzung Pharmacology Semester 5 TablesDocument29 pagesKatzung Pharmacology Semester 5 TablesfatimaPas encore d'évaluation
- Stahl's Essential Psychopharmacology - 5th EdDocument826 pagesStahl's Essential Psychopharmacology - 5th Edjulian.motjeePas encore d'évaluation
- CINP - Antidepressant MedicationsDocument207 pagesCINP - Antidepressant MedicationsAnonymous fPQaCe8Pas encore d'évaluation
- Neurotransmitters Implicated in Psychiatry: MENTOR-Dr. Lokesh Shekhawat Sir. PRESENTOR - Dr. Vibha TomarDocument79 pagesNeurotransmitters Implicated in Psychiatry: MENTOR-Dr. Lokesh Shekhawat Sir. PRESENTOR - Dr. Vibha Tomaranon_870875350100% (1)
- 5 SympatholyticsDocument27 pages5 SympatholyticsHussain RazaPas encore d'évaluation
- Pharmacology 402 February 24, 2010 Mark Hamblin, MD, PHDDocument54 pagesPharmacology 402 February 24, 2010 Mark Hamblin, MD, PHDKarmila Novianti100% (1)
- Concept of PsychobiologyDocument21 pagesConcept of Psychobiologydhivyaram100% (1)
- Synthesis of Adrenaline: Prepared by Sandesh Nair Roll No 204Document20 pagesSynthesis of Adrenaline: Prepared by Sandesh Nair Roll No 204Sreejith Menon0% (1)
- Noradrenaline NeomedDocument3 pagesNoradrenaline NeomedArif212 ArifPas encore d'évaluation
- Pharma Unit 8 - Anti - Adrenergic Drugs - 2 of 2Document21 pagesPharma Unit 8 - Anti - Adrenergic Drugs - 2 of 2MIbrahimPas encore d'évaluation
- Product Development Pipeline - FebruaryDocument4 pagesProduct Development Pipeline - FebruaryCebin VarghesePas encore d'évaluation
- Pharmacology Charts PDFDocument88 pagesPharmacology Charts PDFMohamad Samir90% (10)
- FERNANDO Kristelle C. LectureDocument21 pagesFERNANDO Kristelle C. LectureKervin Jude D. ManahanPas encore d'évaluation
- GPL Test OAT BookDocument40 pagesGPL Test OAT BookJalya JalyaPas encore d'évaluation
- Farma ClinicaDocument43 pagesFarma ClinicaDoina SvetPas encore d'évaluation
- 2023Document25 pages2023yg hanPas encore d'évaluation
- Chemistry MRP Sample For Inspire.Document31 pagesChemistry MRP Sample For Inspire.MridulPas encore d'évaluation
- Investigation of The Effects of Selected Activities Onsecretion of Serotonin Hormone and Some Other HormonesDocument34 pagesInvestigation of The Effects of Selected Activities Onsecretion of Serotonin Hormone and Some Other HormonesGökhan DoğanPas encore d'évaluation
- POLYGRAPHYDocument137 pagesPOLYGRAPHYASUS ZENFONEPas encore d'évaluation
- Ncmb317 Lecture: BSN 3Rd Year 2Nd Semester Prelim 2023: Bachelor of Science in Nursing 3YBDocument62 pagesNcmb317 Lecture: BSN 3Rd Year 2Nd Semester Prelim 2023: Bachelor of Science in Nursing 3YB2 - GUEVARRA, KYLE JOSHUA M.Pas encore d'évaluation
- The Chemistry of Love 1Document3 pagesThe Chemistry of Love 1maudyPas encore d'évaluation
- Bipolar Disorder Concept MapDocument1 pageBipolar Disorder Concept Mapashleydean100% (1)
- Evaluation of Antidepressant Activity of Tramadol and Tramadol Plus Imipramine Using Reserpine Induced Hypothermia Model On Experimental AnimalsDocument6 pagesEvaluation of Antidepressant Activity of Tramadol and Tramadol Plus Imipramine Using Reserpine Induced Hypothermia Model On Experimental AnimalsxiuhtlaltzinPas encore d'évaluation
- The Pharmacology of Adrenergic Receptors PDFDocument15 pagesThe Pharmacology of Adrenergic Receptors PDFKathryn MitchellPas encore d'évaluation
- Hormones of The Adrenal Medulla For LabconDocument57 pagesHormones of The Adrenal Medulla For LabconnikkitingPas encore d'évaluation
- Autonomic DrugsDocument107 pagesAutonomic DrugsMaria Mercedes LeivaPas encore d'évaluation
- Adrenergic ReceptorsDocument9 pagesAdrenergic Receptorsrababmohsin110100% (1)