Vous aimerez peut-être aussi
- Appendectomy During Pregnancy Rates Safety and Outcomes Over A Five Year Period A Hospital Based Follow Up Study - En.esDocument8 pagesAppendectomy During Pregnancy Rates Safety and Outcomes Over A Five Year Period A Hospital Based Follow Up Study - En.estolstoy che rodriguez valderramaPas encore d'évaluation
- BP - OftDocument7 pagesBP - Ofttolstoy che rodriguez valderrama0% (1)
- CASO CLINICO Cancer Endometrio 2Document1 pageCASO CLINICO Cancer Endometrio 2tolstoy che rodriguez valderrama100% (1)
- Otorrino Preguntas CtoDocument4 pagesOtorrino Preguntas Ctotolstoy che rodriguez valderramaPas encore d'évaluation
- NEUROCIRUGIADocument6 pagesNEUROCIRUGIAtolstoy che rodriguez valderramaPas encore d'évaluation
- Cancer de Endometrio Tipo 2Document23 pagesCancer de Endometrio Tipo 2tolstoy che rodriguez valderramaPas encore d'évaluation
- UROLOGÍA DesgPer07Document5 pagesUROLOGÍA DesgPer07Edwin Gustavo Sandoval Torres0% (1)
- Or1 P Test 2VDocument2 pagesOr1 P Test 2Vtolstoy che rodriguez valderramaPas encore d'évaluation
- PsquiatriaDocument48 pagesPsquiatriatolstoy che rodriguez valderramaPas encore d'évaluation
- Banco 6Document11 pagesBanco 6magaPas encore d'évaluation
- OFTALMO DesgPer07Document4 pagesOFTALMO DesgPer07victorPas encore d'évaluation
- Incontinencia-Urinaria ExposicionDocument28 pagesIncontinencia-Urinaria Exposiciontolstoy che rodriguez valderramaPas encore d'évaluation
- Afecciones Del EscrotoDocument16 pagesAfecciones Del Escrototolstoy che rodriguez valderramaPas encore d'évaluation
- Of1 P Test 2VDocument2 pagesOf1 P Test 2Vtolstoy che rodriguez valderramaPas encore d'évaluation
- Transtorno de La Personalidad Obsesivo CompulsivoDocument15 pagesTranstorno de La Personalidad Obsesivo Compulsivotolstoy che rodriguez valderramaPas encore d'évaluation
- NEFROLOGIA Resumen 2018 PDFDocument32 pagesNEFROLOGIA Resumen 2018 PDFArnoldTito100% (1)
- Hiperplasia Benigna PorstaticaDocument13 pagesHiperplasia Benigna Porstaticatolstoy che rodriguez valderramaPas encore d'évaluation
- 006 - Exploración Física Del OidoDocument9 pages006 - Exploración Física Del OidoFonoaudiología UST'Pas encore d'évaluation
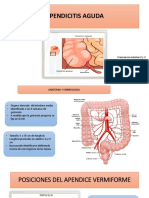
- Apendicitis AgudaDocument15 pagesApendicitis Agudatolstoy che rodriguez valderramaPas encore d'évaluation
- Otitis Media Aguda. Diagnóstico Y Manejo PrácticoDocument9 pagesOtitis Media Aguda. Diagnóstico Y Manejo PrácticoGela AfmaPas encore d'évaluation
- El Crecimiento Económico y La PobrezaDocument1 pageEl Crecimiento Económico y La Pobrezatolstoy che rodriguez valderramaPas encore d'évaluation
- Clase Enam Upao Agosto - 2015nefrologiaDocument360 pagesClase Enam Upao Agosto - 2015nefrologiatolstoy che rodriguez valderramaPas encore d'évaluation
- Conjuntiva PDFDocument44 pagesConjuntiva PDFAldo Negrillo GuzmanPas encore d'évaluation
- PresentacionDocument1 pagePresentaciontolstoy che rodriguez valderramaPas encore d'évaluation
- Becker, Walter - Otorrinolaringologia Manual IlustradoDocument399 pagesBecker, Walter - Otorrinolaringologia Manual IlustradoCarlos Mauricio Ramirez100% (6)
- Test Mini MentalDocument1 pageTest Mini MentalClaudia Toro60% (5)
- Cirugia RoboticaDocument2 pagesCirugia Roboticatolstoy che rodriguez valderramaPas encore d'évaluation
- Banco de PreguntasDocument330 pagesBanco de PreguntasAna CastilloPas encore d'évaluation
- Apendicitis AgudaDocument15 pagesApendicitis Agudatolstoy che rodriguez valderramaPas encore d'évaluation
- NEUMOLOGIA ENAM PRESENCIal 1ra VUELTA PDFDocument245 pagesNEUMOLOGIA ENAM PRESENCIal 1ra VUELTA PDFtolstoy che rodriguez valderramaPas encore d'évaluation
- Tema 3 de Bacteriologia LabDocument10 pagesTema 3 de Bacteriologia Labandrea alzamPas encore d'évaluation
- Membrana CitoplasmáticaDocument56 pagesMembrana CitoplasmáticaGloria Dolores Hernández SolománPas encore d'évaluation
- Guías BIOQUIMICADocument70 pagesGuías BIOQUIMICAYesiid Fernando GuerreroPas encore d'évaluation
- Fundamentos de BacteriologíaDocument181 pagesFundamentos de BacteriologíaGuzvi Dany96% (24)
- Solano 14 - Tecnologia de Altas PresionesDocument26 pagesSolano 14 - Tecnologia de Altas PresionesKelvin Flores SeclenPas encore d'évaluation
- Disposición de Tóxicos. Toxicocinética IDocument16 pagesDisposición de Tóxicos. Toxicocinética IRenatto Taniguchi Salvatierra100% (1)
- Infografia BiologiaDocument2 pagesInfografia BiologiaCarmen CamargoPas encore d'évaluation
- Biología Del Daño y Reparación MuscularDocument7 pagesBiología Del Daño y Reparación MuscularBalam CaballeroPas encore d'évaluation
- Preguntas Leccion PDFDocument20 pagesPreguntas Leccion PDFDaniela AlejandroPas encore d'évaluation
- BiologiaDocument36 pagesBiologiabergPas encore d'évaluation
- Biología - Tarea 2 - Grupo 30176 - 64Document25 pagesBiología - Tarea 2 - Grupo 30176 - 64Ana Lucia CardonaPas encore d'évaluation
- Libro Biol Cel y Ecosist Enero 2020 PDFDocument161 pagesLibro Biol Cel y Ecosist Enero 2020 PDFCarlos Daniel Sarabia CienfuegosPas encore d'évaluation
- Estructura y propiedades de las membranas biológicasDocument5 pagesEstructura y propiedades de las membranas biológicasNayumi MavilaPas encore d'évaluation
- Guía de Problemas N°6 - Bioquímica I 2020 - UNRNDocument3 pagesGuía de Problemas N°6 - Bioquímica I 2020 - UNRNGabii Van CauwenberghePas encore d'évaluation
- Extraccion de Adn CASERODocument3 pagesExtraccion de Adn CASERODiego Gpe Ac AcPas encore d'évaluation
- EXTRACCIÓN DE ADN (Ácido DesoxirribonucleicoDocument2 pagesEXTRACCIÓN DE ADN (Ácido DesoxirribonucleicoEric Bertel SalazarPas encore d'évaluation
- Mapa Mental Membrana PlasmaticaDocument1 pageMapa Mental Membrana PlasmaticaBrian HodzPas encore d'évaluation
- NeurologiaDocument4 pagesNeurologiafioreli pereyraPas encore d'évaluation
- Karp - Potenciales de Membrana (1Document59 pagesKarp - Potenciales de Membrana (1julianPas encore d'évaluation
- Cuestionario y Autoevaluación 7Document5 pagesCuestionario y Autoevaluación 7Nicole EvansPas encore d'évaluation
- Permeabilidad Celular y Fragilidad Osmótica de Los EritrocitosDocument2 pagesPermeabilidad Celular y Fragilidad Osmótica de Los EritrocitosPerlaZavaleta0% (1)
- Mecanismos de ResistenciaDocument8 pagesMecanismos de ResistenciaJohn Maccapa ChancaPas encore d'évaluation
- Investigación Previa ADNDocument2 pagesInvestigación Previa ADNGuadalupe hernandezPas encore d'évaluation
- Membranas MicroporosasDocument2 pagesMembranas MicroporosasyosPas encore d'évaluation
- Lectura Informativa S3Document2 pagesLectura Informativa S3Taly Llarita Guevara CulquiPas encore d'évaluation
- Proyecto CN La Celula VegetalDocument35 pagesProyecto CN La Celula VegetalSuper Erick gamerPas encore d'évaluation
- Industrialización del huevo: proceso de producciónDocument52 pagesIndustrialización del huevo: proceso de producciónDaniel BurgosPas encore d'évaluation
- Gluco CalixDocument10 pagesGluco CalixNikole FreziitaPas encore d'évaluation
- Unidad 1 Procesos Ladbe 2Document81 pagesUnidad 1 Procesos Ladbe 2Sandra Garcia MedinaPas encore d'évaluation
- CElulas Procariota y Eucariota FinalDocument2 pagesCElulas Procariota y Eucariota FinalDeicy Marcela Garcia MoraPas encore d'évaluation