Vous aimerez peut-être aussi
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- Iowa Football 2017 PDFDocument98 pagesIowa Football 2017 PDFKadir Ozcan100% (4)
- David Laid DUP ProgramDocument12 pagesDavid Laid DUP Programshiv kakkar50% (24)
- Rab-RabGAP FTIR X-Ray StudyDocument6 pagesRab-RabGAP FTIR X-Ray StudyKadir OzcanPas encore d'évaluation
- CTE Tau Filaments DISTDocument20 pagesCTE Tau Filaments DISTKadir OzcanPas encore d'évaluation
- Writing The Winter Off Season Program Mike GentryDocument7 pagesWriting The Winter Off Season Program Mike GentryMatt PartneyPas encore d'évaluation
- The Following Physical Performance Testing Standards Have Been Set For The 2014-15 IRB World Sevens SeriesDocument1 pageThe Following Physical Performance Testing Standards Have Been Set For The 2014-15 IRB World Sevens SeriesKadir OzcanPas encore d'évaluation
- Warm-Up and Strength Training EssentialsDocument141 pagesWarm-Up and Strength Training Essentialssjmcconnell100% (2)
- Jefferson CodeofconductDocument20 pagesJefferson CodeofconductKadir OzcanPas encore d'évaluation
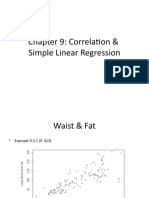
- Waist and Fat RelationshipDocument17 pagesWaist and Fat RelationshipKadir OzcanPas encore d'évaluation
- TDP-43 Gains Function Due To PerturbedDocument20 pagesTDP-43 Gains Function Due To PerturbedKadir OzcanPas encore d'évaluation
- DH Heaviest Weighted Pull-Up Total TemplateDocument2 pagesDH Heaviest Weighted Pull-Up Total TemplateKadir OzcanPas encore d'évaluation
- Bill Star 5X5 CalculatorDocument12 pagesBill Star 5X5 CalculatorRyan PanPas encore d'évaluation
- Boost Install InstructionsDocument2 pagesBoost Install InstructionsKadir OzcanPas encore d'évaluation
- TDP-43 Gains Function Due To PerturbedDocument20 pagesTDP-43 Gains Function Due To PerturbedKadir OzcanPas encore d'évaluation
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (894)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (265)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (119)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Cucumber Extract: A Moisturizing and Antioxidant Ingredient for Skin and Hair CareDocument4 pagesCucumber Extract: A Moisturizing and Antioxidant Ingredient for Skin and Hair CareNarongchai Pongpan100% (1)
- Antiaging Effects of Topical Lactobionic Acid - Results of A Controlled Usage StudyDocument7 pagesAntiaging Effects of Topical Lactobionic Acid - Results of A Controlled Usage StudyLwsynergylab OfficialPas encore d'évaluation
- Foods That Helps Slow Down Aging - Rhuby AbenojaDocument7 pagesFoods That Helps Slow Down Aging - Rhuby AbenojaRHUBY ABENOJAPas encore d'évaluation
- Journal of Medicinal FoodDocument7 pagesJournal of Medicinal FoodАлександр КолядаPas encore d'évaluation
- Advertorial - Sunekos+200 - v.2 LIFTING FACIAL COM HA E AMINOÁCIDOSDocument2 pagesAdvertorial - Sunekos+200 - v.2 LIFTING FACIAL COM HA E AMINOÁCIDOSGustavo Henrique MüllerPas encore d'évaluation
- Flyer Workshop Nanopore Presentation - LowDocument2 pagesFlyer Workshop Nanopore Presentation - LowGulsher AhmedPas encore d'évaluation
- Formulation and Evaluation of Herbal Cream Containing Curcuma LongaDocument7 pagesFormulation and Evaluation of Herbal Cream Containing Curcuma LongaAnonymous uDOOlXxOIAPas encore d'évaluation
- Skin, Hair & Nails Supplement Guide - Examine - 2020 - Examine - Anna's ArchiveDocument75 pagesSkin, Hair & Nails Supplement Guide - Examine - 2020 - Examine - Anna's Archivesimondon11Pas encore d'évaluation
- Nonsurgical Treatments For Infraorbital RejuvenationDocument11 pagesNonsurgical Treatments For Infraorbital RejuvenationHuy Hoàng GiaPas encore d'évaluation
- UV Effects On SkinDocument14 pagesUV Effects On SkinCodrut GeorgescuPas encore d'évaluation
- IPL - Skin Rejuvenation - PROTOCOL PDFDocument45 pagesIPL - Skin Rejuvenation - PROTOCOL PDFmochkurniawan83% (6)
- MERCK - Advance Sun ProtectionDocument59 pagesMERCK - Advance Sun ProtectionCaroline LessaPas encore d'évaluation
- Theresa Beauty 2020 Price List (JE:CL:TPY) - Compressed PDFDocument39 pagesTheresa Beauty 2020 Price List (JE:CL:TPY) - Compressed PDFSandra Koh100% (1)
- Uvinul A PlusDocument11 pagesUvinul A PlusHernanValencia100% (1)
- Phytocosmetics and Cosmetic Science (Nattaya Lourith, Karl Wah-Keung Tsim) (Z-Library)Document170 pagesPhytocosmetics and Cosmetic Science (Nattaya Lourith, Karl Wah-Keung Tsim) (Z-Library)Hector Santiago Lopez AcostaPas encore d'évaluation
- Cosmetic Facial SurgeryDocument785 pagesCosmetic Facial SurgeryStelescu Andras100% (4)
- Chemical Peels - Principles, Peeling Agents, and Pretreatment Assessment - UpToDateDocument16 pagesChemical Peels - Principles, Peeling Agents, and Pretreatment Assessment - UpToDateMichelle Caracas Aponte100% (1)
- Antioxidants 10 01315Document24 pagesAntioxidants 10 01315carloshgmedeirosPas encore d'évaluation
- Plant Polyphenols As Green Sunscreen Ingredients A Systematic ReviewDocument65 pagesPlant Polyphenols As Green Sunscreen Ingredients A Systematic ReviewViey RyzalPas encore d'évaluation
- Valorization of CoffeeDocument9 pagesValorization of CoffeeEditor IJTSRDPas encore d'évaluation
- Feeding The SkinDocument12 pagesFeeding The SkinsunnybeaPas encore d'évaluation
- The Efficacy and Safety of Sunscreen Use For The Prevention of Skin CancerDocument7 pagesThe Efficacy and Safety of Sunscreen Use For The Prevention of Skin CancerBryan NguyenPas encore d'évaluation
- Richa Kothari PDFDocument4 pagesRicha Kothari PDFChinta Ula RanaPas encore d'évaluation
- Assessment of Laser Effects On Skin RejuvenationDocument8 pagesAssessment of Laser Effects On Skin RejuvenationOrel LachianiPas encore d'évaluation
- Vitamin C (CELLEX-C Ascorbic Acid 10%)Document8 pagesVitamin C (CELLEX-C Ascorbic Acid 10%)produksi roiPas encore d'évaluation
- Ingredient For Anti-Pollution PDFDocument131 pagesIngredient For Anti-Pollution PDFAkash SharmaPas encore d'évaluation
- Mesotherapy Poster PDFDocument2 pagesMesotherapy Poster PDFgemm88Pas encore d'évaluation
- Cosmetics: Anti-Aging Properties of Plant Stem Cell ExtractsDocument8 pagesCosmetics: Anti-Aging Properties of Plant Stem Cell ExtractsMohmmad AmannPas encore d'évaluation
- Cuidado de La PielDocument22 pagesCuidado de La PielSummer HornePas encore d'évaluation
- Review Article: Skin Photoaging and The Role of Antioxidants in Its PreventionDocument12 pagesReview Article: Skin Photoaging and The Role of Antioxidants in Its PreventionDessy Erlyani Mugita SariPas encore d'évaluation