Vous aimerez peut-être aussi
- Clasificacion, Etiologia y Epidemiologia Del AcvDocument9 pagesClasificacion, Etiologia y Epidemiologia Del AcvRossio Ortuño LoboPas encore d'évaluation
- Circulacion Por Regiones EspecialesDocument10 pagesCirculacion Por Regiones EspecialesCelvin Rodríguez100% (1)
- FARMACOLOGÍADocument16 pagesFARMACOLOGÍAPaul AlexanderPas encore d'évaluation
- Integración funcional del celador en el área de urgenciasD'EverandIntegración funcional del celador en el área de urgenciasPas encore d'évaluation
- 2lectura de Aga EabDocument4 pages2lectura de Aga EabHector David Rodriguez AquinoPas encore d'évaluation
- Taller de Fármaco Caso 2Document7 pagesTaller de Fármaco Caso 2santiagoPas encore d'évaluation
- Choque de La PuntaDocument9 pagesChoque de La PuntaSandra Panuko100% (1)
- EdemaDocument42 pagesEdemaLaura DominguezPas encore d'évaluation
- Cuadro Comparativo RespiDocument4 pagesCuadro Comparativo RespiHumberto BecePas encore d'évaluation
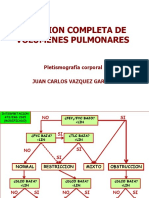
- Medicion Completa de Volumenes PulmonaresDocument95 pagesMedicion Completa de Volumenes PulmonaresmavhelllinaPas encore d'évaluation
- Árbol BronquialDocument6 pagesÁrbol BronquialIsabel Fernandez BancesPas encore d'évaluation
- 004 Transporte de Oxigeno y Dioxido de CarbonoDocument32 pages004 Transporte de Oxigeno y Dioxido de CarbonoThulio MoutinhoPas encore d'évaluation
- Gauna CompletoDocument221 pagesGauna CompletoMatheus MatrealPas encore d'évaluation
- Medicamentos de Una FarmaciaDocument12 pagesMedicamentos de Una FarmaciaIngridPas encore d'évaluation
- Capitulo 17 Examen Del AbdomenDocument38 pagesCapitulo 17 Examen Del AbdomenLuisdeLeonPas encore d'évaluation
- Cateter Swan GanzDocument25 pagesCateter Swan GanzCarolina Herrera HincapiePas encore d'évaluation
- Gases ArterialesDocument6 pagesGases Arterialesapolion00Pas encore d'évaluation
- Síndrome Parasitario PresentacionDocument35 pagesSíndrome Parasitario PresentacionIsmael NavarroPas encore d'évaluation
- Fisiopatologia Respiratoria (Trastornos de Difusion)Document30 pagesFisiopatologia Respiratoria (Trastornos de Difusion)Roger Alejandro MontoyaPas encore d'évaluation
- Ventilacion y Espacio MuertoDocument10 pagesVentilacion y Espacio Muertoainsaurralde_1Pas encore d'évaluation
- Atención de Enfermería A Pacientes Con Afecciones Del Aparato CardiovascularDocument13 pagesAtención de Enfermería A Pacientes Con Afecciones Del Aparato CardiovascularLibia Castellanos100% (1)
- Fármacos Antihipertensivos OralesDocument41 pagesFármacos Antihipertensivos OralesPaul Mendoza SalasPas encore d'évaluation
- Equilibrio Acido BaseDocument9 pagesEquilibrio Acido BaseJavier ToalaPas encore d'évaluation
- Angina Inestable e Infarto Sin Elevacion Del Segmento STDocument94 pagesAngina Inestable e Infarto Sin Elevacion Del Segmento STAlex CarranzaPas encore d'évaluation
- Guia Patologia Pulmon y Pleura (Robbins y Cotran)Document3 pagesGuia Patologia Pulmon y Pleura (Robbins y Cotran)Jeick Martínez100% (1)
- Generalidades de OxigenoterapiaDocument31 pagesGeneralidades de Oxigenoterapiaduvan figueroa100% (1)
- CASOS CLINICOS GASTROINTESTINAL 2r.Document7 pagesCASOS CLINICOS GASTROINTESTINAL 2r.Claudia Lopez LozaPas encore d'évaluation
- Insuficiencia Periferica Arterial y VenosaDocument37 pagesInsuficiencia Periferica Arterial y VenosaDany Antezana CanoPas encore d'évaluation
- Bases Fisicas de La ElectrocardiografiaDocument47 pagesBases Fisicas de La Electrocardiografiamikichiko2349Pas encore d'évaluation
- Electrocardiograma PDFDocument4 pagesElectrocardiograma PDFMayrelis BohorquezPas encore d'évaluation
- Fisiologia - Mecanica Ventilatoria y Curvas EspirometricasDocument63 pagesFisiologia - Mecanica Ventilatoria y Curvas EspirometricasUSMP FN ARCHIVOSPas encore d'évaluation
- Embriologia de Corazon PDFDocument10 pagesEmbriologia de Corazon PDFFrancisca Fernanda NorambuenaPas encore d'évaluation
- Examen de MorfofisiologiaDocument2 pagesExamen de MorfofisiologiaMike0% (1)
- Anticuerpos y AntigenosDocument28 pagesAnticuerpos y Antigenoswaldrapa29Pas encore d'évaluation
- Staphylococcus Aureus - Bacteria Patogena PDFDocument7 pagesStaphylococcus Aureus - Bacteria Patogena PDFCarlos Sierra LandinPas encore d'évaluation
- Metabolismo de Quilomicrones y VLDLDocument3 pagesMetabolismo de Quilomicrones y VLDLCRISTINA DIAZ LOPEZPas encore d'évaluation
- CASO CLINICO HipertiroidismoDocument3 pagesCASO CLINICO Hipertiroidismobrighit wendy sandoval alayoPas encore d'évaluation
- Electrocardiograma 2021Document13 pagesElectrocardiograma 2021csos csos0% (1)
- Taller Hipotálamo HipófisisDocument3 pagesTaller Hipotálamo HipófisisNoah Annelise OrtizPas encore d'évaluation
- Triangulo de CalotDocument6 pagesTriangulo de CalotDaisuke0% (1)
- 2.corazon Como BombaDocument8 pages2.corazon Como BombaDaniela AbadPas encore d'évaluation
- Gastrosquisis OnfaloceleDocument19 pagesGastrosquisis OnfaloceleValeriaPas encore d'évaluation
- Osteonecrosis, Osteocondritis y OsteocondrosisDocument7 pagesOsteonecrosis, Osteocondritis y OsteocondrosisCambriaChicoPas encore d'évaluation
- Hipertensión Intracraneal - Leonardo CruchagaDocument23 pagesHipertensión Intracraneal - Leonardo CruchagaMa Del Pilar FerraPas encore d'évaluation
- Factores de Riesgo Cardiovascular - Ángel González PDFDocument236 pagesFactores de Riesgo Cardiovascular - Ángel González PDFOmar Hernandez RodriguezPas encore d'évaluation
- Embriologia de ToraxDocument4 pagesEmbriologia de Toraxapi-19734073Pas encore d'évaluation
- Tarea Sistema Endocrino 1Document7 pagesTarea Sistema Endocrino 1Fernando BolivaristaPas encore d'évaluation
- Aparato UrinarioDocument34 pagesAparato UrinarioEstela Castro MontesPas encore d'évaluation
- Monitoreo No InvasivoDocument29 pagesMonitoreo No InvasivoDr. Jorge Alam50% (2)
- Sindromes Pleuropulmonares 1Document9 pagesSindromes Pleuropulmonares 1MAYOPas encore d'évaluation
- Potencial de Accion Del Musculo Cardiaco y EkgDocument43 pagesPotencial de Accion Del Musculo Cardiaco y EkgPaola Custodio GonzalesPas encore d'évaluation
- HiponatremiaDocument10 pagesHiponatremiaGabyflobePas encore d'évaluation
- Oclusión Intestinal, Acidosis Metabólica, Deshidratación GraveDocument5 pagesOclusión Intestinal, Acidosis Metabólica, Deshidratación GraveSadie Elitzel Aguilar50% (2)
- Alteraciones Acido BaseDocument37 pagesAlteraciones Acido BaseClareth De Horta CadenaPas encore d'évaluation
- Clasificación Palm CoeinDocument4 pagesClasificación Palm Coeinfelipe sanchezPas encore d'évaluation
- Módulo de Principios de Epidemiología para El Control de Enfermedades (Organización Panamericana de La Salud)Document478 pagesMódulo de Principios de Epidemiología para El Control de Enfermedades (Organización Panamericana de La Salud)Linette Lagunes LaraPas encore d'évaluation
- Capitulo 5 BaynesDocument7 pagesCapitulo 5 BaynesClaudio Cofré MansillaPas encore d'évaluation
- Marketing 4.0Document3 pagesMarketing 4.0Teresa Consuelo Espinoza Saldaña100% (1)
- RofDocument45 pagesRofTeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- Preguntas de MacroeconomíaDocument5 pagesPreguntas de MacroeconomíaTeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- Preguntas de MacroeconomíaDocument5 pagesPreguntas de MacroeconomíaTeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- Plan - 13771 - Manual de Organizacion y Funciones (Parte5) - 2009 PDFDocument29 pagesPlan - 13771 - Manual de Organizacion y Funciones (Parte5) - 2009 PDFDavid RoblesPas encore d'évaluation
- Preguntas de MacroeconomíaDocument5 pagesPreguntas de MacroeconomíaTeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- La Procesión de Las ÁnimasDocument2 pagesLa Procesión de Las ÁnimasTeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- MICROECONOMIADocument1 pageMICROECONOMIATeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- Psicologia Del LogroDocument10 pagesPsicologia Del LogroTeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- ExitoyutopiaDocument20 pagesExitoyutopiaTeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- Principios de La Contribucion MarginalDocument31 pagesPrincipios de La Contribucion MarginalTeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- El Papel y La Importancia de Las CelulasDocument4 pagesEl Papel y La Importancia de Las CelulasTeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- Principales Bloques EconómicosDocument3 pagesPrincipales Bloques EconómicosTeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- BambasDocument35 pagesBambasTeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- Qué Es Un Movie MakerDocument3 pagesQué Es Un Movie MakerTeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- Definicion de La Responsabilidad Social EmpresarialDocument2 pagesDefinicion de La Responsabilidad Social EmpresarialTeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- Capitulo V EconomiaDocument11 pagesCapitulo V EconomiaTeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- La Etica Kantiana Libro SingerDocument12 pagesLa Etica Kantiana Libro SingerTeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- Tipos de Empresa MetodologiaDocument1 pageTipos de Empresa MetodologiaTeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- Responsabilidad Social - Grupal de La Empresa GloriaDocument19 pagesResponsabilidad Social - Grupal de La Empresa GloriaYessi Antunez Condezo25% (4)
- OsificaciónDocument14 pagesOsificaciónTeresa Consuelo Espinoza Saldaña100% (2)
- Malformaciones Del ÚteroDocument19 pagesMalformaciones Del ÚteroTeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- Etica KantianaDocument8 pagesEtica KantianaPavel Gabriel Tapia RomeroPas encore d'évaluation
- Acto JuridicoDocument11 pagesActo JuridicoTeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- CardiovascularDocument9 pagesCardiovascularTeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- Estimulación Ovárica y Extracción de OvocitosDocument8 pagesEstimulación Ovárica y Extracción de OvocitosTeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- Embrio Sem 3Document6 pagesEmbrio Sem 3Teresa Consuelo Espinoza SaldañaPas encore d'évaluation
- Histo Sem 3 Introducción y ConclusionesDocument4 pagesHisto Sem 3 Introducción y ConclusionesTeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- Anomalías Cromosómicas SexualesDocument10 pagesAnomalías Cromosómicas SexualesTeresa Consuelo Espinoza SaldañaPas encore d'évaluation
- Apuntes Tema 5Document15 pagesApuntes Tema 5m.iranzo.g.14Pas encore d'évaluation
- Biologia Desarrollo ResumenDocument4 pagesBiologia Desarrollo ResumenFidel CocónPas encore d'évaluation
- Den Ti NoDocument26 pagesDen Ti NoJacquelinePas encore d'évaluation
- Respuestas Celulares Al Estrés y Las Agresiones TóxicasDocument62 pagesRespuestas Celulares Al Estrés y Las Agresiones TóxicasElena González CruzPas encore d'évaluation
- GUIAS Sexto 1 PERIODO BIOLOGIADocument10 pagesGUIAS Sexto 1 PERIODO BIOLOGIALuis Alexander Torres MartínezPas encore d'évaluation
- Simulacro 01 080922Document15 pagesSimulacro 01 080922Jhoanmy CalixtoPas encore d'évaluation
- Ultrasonido UNeBDocument61 pagesUltrasonido UNeBpfuentes90Pas encore d'évaluation
- Centrifugación Zonal y UltracentrifugaciónDocument2 pagesCentrifugación Zonal y UltracentrifugaciónGABRIELA ARAGON SERRANOPas encore d'évaluation
- Planificacion CCNN 2022 OctavoDocument6 pagesPlanificacion CCNN 2022 OctavoNathaly Torres0% (1)
- TEORIA 2 Estructura BacterianaDocument39 pagesTEORIA 2 Estructura BacterianajackridenPas encore d'évaluation
- La CélulaDocument8 pagesLa CélulaAngelinne Garcia APas encore d'évaluation
- 70 Preguntas Con Respuesta - Introducción A La FisiologíaDocument9 pages70 Preguntas Con Respuesta - Introducción A La FisiologíaMarrrrrPas encore d'évaluation
- Historia Completa Del ScioDocument31 pagesHistoria Completa Del ScioFacundo AllendePas encore d'évaluation
- Tesis-Cushuro FinalDocument43 pagesTesis-Cushuro Finalshadow dark100% (1)
- Examen de Fisiología CelularDocument5 pagesExamen de Fisiología CelularJavier GM0% (1)
- Alta Presion HidrostaticaDocument19 pagesAlta Presion HidrostaticamemolisisPas encore d'évaluation
- Glosario de TerminosDocument6 pagesGlosario de TerminosSaraGarciaPas encore d'évaluation
- 3 Guía Tercerp.a. Origen de La VidaDocument4 pages3 Guía Tercerp.a. Origen de La VidaDiego CelyPas encore d'évaluation
- Taxonomia de Los VirusDocument75 pagesTaxonomia de Los VirusSonHyunFelixPas encore d'évaluation
- Resumen2 Psicología Del Desarrollo - El Ciclo Vital. J.W.SantrockDocument15 pagesResumen2 Psicología Del Desarrollo - El Ciclo Vital. J.W.SantrockRaquel MarlascaPas encore d'évaluation
- Clasificacion BacterianaDocument20 pagesClasificacion BacterianaBrenda Dalila Barrientos MartinezPas encore d'évaluation
- La Glandula PinealDocument15 pagesLa Glandula PinealCris CavasottoPas encore d'évaluation
- Metabolismo 1 7Document7 pagesMetabolismo 1 7Lizeth PinoPas encore d'évaluation
- Eval Diag 1° DifesoDocument3 pagesEval Diag 1° DifesoCarlos TorresPas encore d'évaluation
- Biologia TTTDocument2 pagesBiologia TTTDiaz PipePas encore d'évaluation
- Examen Biolog°a Primer Trimestre RESPUESTASDocument2 pagesExamen Biolog°a Primer Trimestre RESPUESTASjonathanPas encore d'évaluation
- Parcial 1 Laboratorio BiomolDocument11 pagesParcial 1 Laboratorio BiomolSebastianAlvarezAtehortuaPas encore d'évaluation
- Tratamientos AnticancerDocument133 pagesTratamientos AnticancerLeonidas737Pas encore d'évaluation
- Celula, Agua y PHDocument39 pagesCelula, Agua y PHDaniel MirandaPas encore d'évaluation
- Sesion de Aprendizaje El Origen de La VidaDocument4 pagesSesion de Aprendizaje El Origen de La VidamcozlastraPas encore d'évaluation