Vous aimerez peut-être aussi
- New Debate Human Embryo: A Biological DefinitionDocument7 pagesNew Debate Human Embryo: A Biological DefinitionPetru CernatPas encore d'évaluation
- Bioethics B1B017025 Syamsul Ma'arif Short Paper CloneDocument3 pagesBioethics B1B017025 Syamsul Ma'arif Short Paper CloneSyamsul Ma'arifPas encore d'évaluation
- Stem Cell Presentation Revised September 2015Document28 pagesStem Cell Presentation Revised September 2015nazım tekerPas encore d'évaluation
- Talking Point: Embryonic Human PersonsDocument6 pagesTalking Point: Embryonic Human PersonsheyimherePas encore d'évaluation
- From Stem Cells To IPS Cells. A Passionate JourneyDocument9 pagesFrom Stem Cells To IPS Cells. A Passionate JourneyP Rigoberto CanoPas encore d'évaluation
- Molecular Basis of Organism Differentiation at Cell LevelDocument16 pagesMolecular Basis of Organism Differentiation at Cell Levelkumar siddhantPas encore d'évaluation
- Cloning HumansDocument33 pagesCloning HumansKristine Rhoda Matrido-DolarPas encore d'évaluation
- Stem Cells - overview: Embryonic stem cell therapy for spinal cord injury faces delaysDocument39 pagesStem Cells - overview: Embryonic stem cell therapy for spinal cord injury faces delaysubaidu dikkoPas encore d'évaluation
- The Veterinary Journal: V. Hall, K. Hinrichs, G. Lazzari, D.H. Betts, P. HyttelDocument15 pagesThe Veterinary Journal: V. Hall, K. Hinrichs, G. Lazzari, D.H. Betts, P. HyttelJuniClaudia13Pas encore d'évaluation
- Animal Reproductive Systems: © Cengage Learning 2016. All Rights ReservedDocument41 pagesAnimal Reproductive Systems: © Cengage Learning 2016. All Rights ReservedbutterhaezelPas encore d'évaluation
- Science Aax7890 PDFDocument9 pagesScience Aax7890 PDFAndoingPas encore d'évaluation
- Developmental and Reproductive Biology GuideDocument3 pagesDevelopmental and Reproductive Biology GuideTrxsee RoxasPas encore d'évaluation
- Class X Science Term 2 ReproductionDocument11 pagesClass X Science Term 2 ReproductionMokshita JainPas encore d'évaluation
- HiiDocument19 pagesHiiTruptimayiGiriPas encore d'évaluation
- Introduction to Biomedical ScienceDocument86 pagesIntroduction to Biomedical ScienceBiology BảoPas encore d'évaluation
- Developmental Biology: Fundamental Processes of DevelopmentDocument9 pagesDevelopmental Biology: Fundamental Processes of DevelopmentManisha BishtPas encore d'évaluation
- Define The FollowingDocument19 pagesDefine The FollowingAbhinav AtrayPas encore d'évaluation
- La Clonación.Document15 pagesLa Clonación.Javier Repizo OlayaPas encore d'évaluation
- 2021 orvietoDocument5 pages2021 orvieto99ornelaPas encore d'évaluation
- Bioteknologi 110-Stem CellsDocument14 pagesBioteknologi 110-Stem CellsNahla FaridaPas encore d'évaluation
- Cloning: Examples of Animal Cloning MethodsDocument8 pagesCloning: Examples of Animal Cloning MethodsZuza PodleśnaPas encore d'évaluation
- Testimony Stem CellsDocument9 pagesTestimony Stem CellsKathryn MayPas encore d'évaluation
- Embryo culture: can we perform better than natureDocument17 pagesEmbryo culture: can we perform better than natureHector Salas GarcíaPas encore d'évaluation
- Animal Reproduction & Development: AP BiologyDocument22 pagesAnimal Reproduction & Development: AP Biologyjulie rainesPas encore d'évaluation
- Lecture 7. Basic Developmental Biology and BiotecnologyDocument48 pagesLecture 7. Basic Developmental Biology and BiotecnologyNatsuki HajimePas encore d'évaluation
- Human Reproduction ReviewDocument12 pagesHuman Reproduction ReviewMokYikLamPas encore d'évaluation
- Beginning of LifeDocument25 pagesBeginning of LifeLEEPas encore d'évaluation
- Stem Cell Research Status and EthicsDocument63 pagesStem Cell Research Status and EthicsPoonkodi Tp100% (1)
- Version On-Line ISSN 0717-9502: International Journal of MorphologyDocument8 pagesVersion On-Line ISSN 0717-9502: International Journal of MorphologyMaría AlejandraPas encore d'évaluation
- 10.5 Whole Organism Cloning in Animals COMPLETEDDocument4 pages10.5 Whole Organism Cloning in Animals COMPLETEDhixpatchPas encore d'évaluation
- Embryo Splitting: Middle East Fertility Society JournalDocument7 pagesEmbryo Splitting: Middle East Fertility Society Journal22. Lioni Pasaribu [XA5]Pas encore d'évaluation
- Gene CloningDocument18 pagesGene CloningRatnesh Raman Pathak100% (1)
- Stem Cell (3), BB-Ida PDFDocument50 pagesStem Cell (3), BB-Ida PDFWynona RamliPas encore d'évaluation
- Last-minute Biology revision guide for Class XII students in Jaipur regionDocument18 pagesLast-minute Biology revision guide for Class XII students in Jaipur regionghjPas encore d'évaluation
- Zygote - WikipediaDocument17 pagesZygote - WikipediaBashiir NuurPas encore d'évaluation
- Human Reproductive Endocrinology: Biol/Bmsc 114Document27 pagesHuman Reproductive Endocrinology: Biol/Bmsc 114Marcus EbertPas encore d'évaluation
- Definition:: The Technique of Producing A Genetically Identical Copy of An OrganismDocument44 pagesDefinition:: The Technique of Producing A Genetically Identical Copy of An OrganismmainehoonaPas encore d'évaluation
- BIO 325 Exam 1 ReviewDocument17 pagesBIO 325 Exam 1 ReviewNerdy Notes Inc.Pas encore d'évaluation
- Animal CloningDocument33 pagesAnimal CloningAndi RohandiPas encore d'évaluation
- Kloning Human...Document10 pagesKloning Human...dwi rositaPas encore d'évaluation
- Animal CloningDocument13 pagesAnimal Cloningdebanjana2009Pas encore d'évaluation
- Introduction:-: Applications of Human CloningDocument13 pagesIntroduction:-: Applications of Human CloningGladys PriyaPas encore d'évaluation
- (Zhou Et Al., 2016) Complete Meiosis From Embryonic Stem Cell-Derived Germ Cells in VitroDocument12 pages(Zhou Et Al., 2016) Complete Meiosis From Embryonic Stem Cell-Derived Germ Cells in VitroJulio dR AltavasPas encore d'évaluation
- Embryology: Department of Anatomy & Histology, Khulna Agricultural UniversityDocument42 pagesEmbryology: Department of Anatomy & Histology, Khulna Agricultural Universityআহানাফ তাহমিদ শব্দPas encore d'évaluation
- Ch-9 Reproduction in AnimalsDocument2 pagesCh-9 Reproduction in AnimalsShreya KashyapPas encore d'évaluation
- Study of Normal and Abnormal Prenatal DevelopmentDocument5 pagesStudy of Normal and Abnormal Prenatal DevelopmentPamela MolinaPas encore d'évaluation
- Larsens Human Embryology 4th Edition Schoenwolf Test BankDocument4 pagesLarsens Human Embryology 4th Edition Schoenwolf Test BankJoshuaKnappkmzpc100% (18)
- Biotechnology Introduction: Cloning Techniques and ApplicationsDocument5 pagesBiotechnology Introduction: Cloning Techniques and ApplicationsAfrin IbrahimPas encore d'évaluation
- Human Cloning and Genetic ModificationDocument6 pagesHuman Cloning and Genetic ModificationaballacarlosjasonPas encore d'évaluation
- Pigs ProteomicsDocument7 pagesPigs Proteomicsisabel velezPas encore d'évaluation
- Reprod Medicine Biology - 2004 - NAKAJO - Physical and Mental Development of Children After in Vitro Fertilization andDocument5 pagesReprod Medicine Biology - 2004 - NAKAJO - Physical and Mental Development of Children After in Vitro Fertilization andTirbuAndreiPas encore d'évaluation
- Reproductive Biology and Endocrinology: Oogenesis in Cultures Derived From Adult Human OvariesDocument13 pagesReproductive Biology and Endocrinology: Oogenesis in Cultures Derived From Adult Human Ovaries05-OB-HU-DINA MERCEDES CAMPOSANO SALAZARPas encore d'évaluation
- Gap 055Document15 pagesGap 05505-OB-HU-DINA MERCEDES CAMPOSANO SALAZARPas encore d'évaluation
- Human cloning: An ethical debateDocument4 pagesHuman cloning: An ethical debateXangotPas encore d'évaluation
- CLONING TECHNOLOGYDocument25 pagesCLONING TECHNOLOGYMark Christian M. GonzagaPas encore d'évaluation
- Potential Uses and Ethical Debates of Stem Cell ResearchDocument37 pagesPotential Uses and Ethical Debates of Stem Cell ResearchJesha LibreaPas encore d'évaluation
- Oocyte Control of Ovarian Follicular Development and Function in MammalsDocument10 pagesOocyte Control of Ovarian Follicular Development and Function in MammalsArDi EcHaPas encore d'évaluation
- Live Birth After Transfer of A Single Euploid Vitrified-Warmed Blastocyst According To Standard Timing vs. Timing As Recommended by Endometrial Receptivity AnalysisDocument8 pagesLive Birth After Transfer of A Single Euploid Vitrified-Warmed Blastocyst According To Standard Timing vs. Timing As Recommended by Endometrial Receptivity AnalysisAnh Vũ Hồ NgọcPas encore d'évaluation
- JurnalDocument13 pagesJurnalparkc6922Pas encore d'évaluation
- Pancreatic and Periampullary Carcinoma (Nonendocrine) : Host FactorsDocument26 pagesPancreatic and Periampullary Carcinoma (Nonendocrine) : Host Factorsjuan jose velascoPas encore d'évaluation
- The Influence of KantDocument5 pagesThe Influence of Kantjuan jose velascoPas encore d'évaluation
- Report OMS 2001 PDFDocument404 pagesReport OMS 2001 PDFjuan jose velascoPas encore d'évaluation
- A Guide For Patients: Ge and ErtilityDocument20 pagesA Guide For Patients: Ge and ErtilityDeidre VilleneuvePas encore d'évaluation
- The Surrogacy (Regulation) Act, 2021: Reliminary EctionsDocument21 pagesThe Surrogacy (Regulation) Act, 2021: Reliminary EctionsJay TiwariPas encore d'évaluation
- BIOLOGY Form 5 Chapter 6Document21 pagesBIOLOGY Form 5 Chapter 6Shephard Png100% (3)
- Science Form 2Document19 pagesScience Form 2Mohamad TarmiziPas encore d'évaluation
- Female Reproductive System: Related Kidshealth LinksDocument7 pagesFemale Reproductive System: Related Kidshealth Linksrichy0% (1)
- 12th Bio-Zoology (EM) 2022-2023 SampleDocument17 pages12th Bio-Zoology (EM) 2022-2023 SampleReeshi GunaPas encore d'évaluation
- Anther Pollen Ovule Culture TechniquesDocument11 pagesAnther Pollen Ovule Culture TechniquesVARTIKA KHATI100% (1)
- ZIFT FinalDocument11 pagesZIFT Finalblese56100% (1)
- Lenv62012 Exam WDocument24 pagesLenv62012 Exam Wcarlapaulavicente3774Pas encore d'évaluation
- Annexure 'G': Syllabus BIOLOGY (044) CLASS-XII (2013-2014)Document6 pagesAnnexure 'G': Syllabus BIOLOGY (044) CLASS-XII (2013-2014)shubhamchopra97Pas encore d'évaluation
- Pollen-Pistil Interaction: Pollen Attachment and HydrationDocument6 pagesPollen-Pistil Interaction: Pollen Attachment and HydrationSreeja Raj100% (1)
- Bio IDocument234 pagesBio ISimiYadavPas encore d'évaluation
- A Semi Detailed Lesson Plan in ScienceDocument3 pagesA Semi Detailed Lesson Plan in ScienceI AM ANONYMOUSPas encore d'évaluation
- School Hda. Conchita National High School Grade Level 11 Earth and LifeDocument6 pagesSchool Hda. Conchita National High School Grade Level 11 Earth and LifeCherry Pink VillanuevaPas encore d'évaluation
- Biology Mapping GuideDocument28 pagesBiology Mapping GuideGazar100% (1)
- Xii Biology Assertion-And-ReasoningDocument18 pagesXii Biology Assertion-And-ReasoningKiran RaoPas encore d'évaluation
- Syllabus Class Xi 2019 20Document31 pagesSyllabus Class Xi 2019 20Aniket BasuPas encore d'évaluation
- Lecture On Transgenic AnimalsDocument25 pagesLecture On Transgenic AnimalsMuhammad Ikram Anwar100% (3)
- REPRODUCTION-WPS OfficeDocument6 pagesREPRODUCTION-WPS OfficeGwzwwgPas encore d'évaluation
- Sci6-Q2-Module-7 EditedDocument28 pagesSci6-Q2-Module-7 Editededelberto100% (1)
- Essential guide to oral communicationDocument14 pagesEssential guide to oral communicationarchie bacudPas encore d'évaluation
- Vertebrate Embryology - Randel PDFDocument399 pagesVertebrate Embryology - Randel PDFrandelPas encore d'évaluation
- Class 8 - Science - Reproduction in AnimalsDocument6 pagesClass 8 - Science - Reproduction in AnimalsLucky MishraPas encore d'évaluation
- Lycopodium Habit and Morphology GuideDocument73 pagesLycopodium Habit and Morphology GuideAnjaliPas encore d'évaluation
- Demonstrate Knowledge of The Stages of Child DevelopmentDocument64 pagesDemonstrate Knowledge of The Stages of Child DevelopmentAvaPas encore d'évaluation
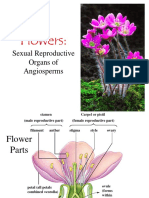
- Flowers: Sexual Reproduction in AngiospermsDocument106 pagesFlowers: Sexual Reproduction in AngiospermsAlfonso PlantillaPas encore d'évaluation
- (Code No. 044) : Term - IDocument6 pages(Code No. 044) : Term - IA KumarPas encore d'évaluation
- Animal Reproduction of Representative Animals ReviewerDocument3 pagesAnimal Reproduction of Representative Animals ReviewerRizza Mel GalleroPas encore d'évaluation
- Instruction On Respect For Human LifeDocument21 pagesInstruction On Respect For Human LifeJBSfanPas encore d'évaluation
- LIFS 1902 - General Biology II: Video 4 - Human DevelopmentDocument20 pagesLIFS 1902 - General Biology II: Video 4 - Human DevelopmentGary TsangPas encore d'évaluation
- Topical Questions Form 3 - BiologyDocument32 pagesTopical Questions Form 3 - BiologyBadmind Jnr100% (2)