Vous aimerez peut-être aussi
- Inmunofarmacología de Los ProbióticosDocument7 pagesInmunofarmacología de Los ProbióticosJosé MaríaPas encore d'évaluation
- Enfermedad de CrohnDocument3 pagesEnfermedad de CrohnBrye IeroPas encore d'évaluation
- Exposicion EIIDocument5 pagesExposicion EIIJavier GomezPas encore d'évaluation
- Candida Albicans Es Una Levadura Comensal Que Reside en Las Membranas Mucosas de Las Cavidades Oral y VaginalDocument4 pagesCandida Albicans Es Una Levadura Comensal Que Reside en Las Membranas Mucosas de Las Cavidades Oral y VaginalDaniel CastilloPas encore d'évaluation
- Candida Albicans Es Una Levadura Comensal Que Reside en Las Membranas Mucosas de Las Cavidades Oral y VaginalDocument4 pagesCandida Albicans Es Una Levadura Comensal Que Reside en Las Membranas Mucosas de Las Cavidades Oral y VaginalDaniel CastilloPas encore d'évaluation
- Immunity in The Gut (Inmunidad en El Intestino)Document2 pagesImmunity in The Gut (Inmunidad en El Intestino)QUIERO18981Pas encore d'évaluation
- Patogenia y Relación HospederoDocument3 pagesPatogenia y Relación HospederoMelissa CelisPas encore d'évaluation
- Microbioma y Enfermedades AlergicasDocument25 pagesMicrobioma y Enfermedades AlergicasTatiana Cáceres HernándezPas encore d'évaluation
- INTERLEUCINAS (Resumen 3)Document5 pagesINTERLEUCINAS (Resumen 3)Kike LópezPas encore d'évaluation
- Tuberculosis ResumenDocument5 pagesTuberculosis ResumenOmar AdalidPas encore d'évaluation
- Maylin Pi NeumoniaDocument6 pagesMaylin Pi Neumoniayvasquezp191Pas encore d'évaluation
- CÉLULAS Y MOLÉCULAS EFECTORAS-resumenDocument1 pageCÉLULAS Y MOLÉCULAS EFECTORAS-resumenLUCIA DEL PILAR LIMACHI ACOSTAPas encore d'évaluation
- Respuesta Inmunológica Frente A HongosDocument2 pagesRespuesta Inmunológica Frente A HongosAlondra VegaPas encore d'évaluation
- CITOKINASDocument2 pagesCITOKINASAnonymous f2hnaTPas encore d'évaluation
- IntroducciónDocument20 pagesIntroducciónMauri GuañunaPas encore d'évaluation
- Inmunomoduladores en PreeclampsiaDocument13 pagesInmunomoduladores en PreeclampsiaErick GaliciaPas encore d'évaluation
- Inmunologia de LepraDocument8 pagesInmunologia de LepraEfrain Teran CarbonoPas encore d'évaluation
- Cómo Actúa El Sistema Inmune Frente A Los Parásitos Helmínti22445Document7 pagesCómo Actúa El Sistema Inmune Frente A Los Parásitos Helmínti22445Jovana CarbajalPas encore d'évaluation
- Tsa3-Ordoñez Vela Leidy StehfanyDocument7 pagesTsa3-Ordoñez Vela Leidy StehfanyLeidy Sthefany Ordoñez VelaPas encore d'évaluation
- MCROBIOTADocument7 pagesMCROBIOTAMarlyn Vanessa Marin RuedaPas encore d'évaluation
- Becerril 4a c03 Parasitologia DigestivoDocument4 pagesBecerril 4a c03 Parasitologia DigestivoRuth Lisbet Chara Chara100% (1)
- Microbiota IntestinalDocument2 pagesMicrobiota Intestinalfernandop.ugunaaPas encore d'évaluation
- Cuarto Corte InmunoDocument86 pagesCuarto Corte InmunoVALENTINA ESCOBAR QUINTEROPas encore d'évaluation
- Tratamiento de La Flora Intestinal Del HombreDocument9 pagesTratamiento de La Flora Intestinal Del HombreDr. corazónPas encore d'évaluation
- FTSistema InmuneDocument14 pagesFTSistema InmuneCanis MajorisPas encore d'évaluation
- H PyloriDocument3 pagesH PyloriCarlos VásquezPas encore d'évaluation
- Adaptaciones ParasitariasDocument4 pagesAdaptaciones ParasitariascriznahpPas encore d'évaluation
- Sistema Inmune MucosasDocument7 pagesSistema Inmune Mucosasabelhenarejos-1Pas encore d'évaluation
- Sindrome de Aspiracion de Liquido Meconial: Ciro Juan Atauje TrilloDocument53 pagesSindrome de Aspiracion de Liquido Meconial: Ciro Juan Atauje TrilloLido Cardenas VargasPas encore d'évaluation
- Respuestas Inmunes Innatas y Adaptativas en El Tracto UrinarioDocument10 pagesRespuestas Inmunes Innatas y Adaptativas en El Tracto Urinarioana reyna100% (2)
- A Resposta Inmunitaria Frente A ParásitosDocument5 pagesA Resposta Inmunitaria Frente A Parásitoscolordeestrellas92Pas encore d'évaluation
- 8 Inmunidad Frente A Bacterias, Virus, Parasitos, Protozoos y ParazoosDocument29 pages8 Inmunidad Frente A Bacterias, Virus, Parasitos, Protozoos y ParazoosFlora Valencia ManzanaresPas encore d'évaluation
- InmunologíaDocument10 pagesInmunologíaevangelinePas encore d'évaluation
- Inmunoestimulantes Orales en PediatriaDocument159 pagesInmunoestimulantes Orales en PediatriaHenry Arista50% (2)
- Presentación Antigénica para El Virus de Influenza.Document3 pagesPresentación Antigénica para El Virus de Influenza.Hernandez MariaPas encore d'évaluation
- Infeccion H Pylori Libro SCGDocument14 pagesInfeccion H Pylori Libro SCGHugo GonzálezPas encore d'évaluation
- Articulo Lepra th1 y th2 PDFDocument15 pagesArticulo Lepra th1 y th2 PDFAndres Felipe ChicaPas encore d'évaluation
- Enteritis Linfoplasmocítica CoralDocument10 pagesEnteritis Linfoplasmocítica Coralmidorino5Pas encore d'évaluation
- Probioticos para La Inmunidad y La Funcion LipidicaDocument16 pagesProbioticos para La Inmunidad y La Funcion LipidicaJulio CorreaPas encore d'évaluation
- Alimentos AntiinflamatoriosDocument16 pagesAlimentos AntiinflamatoriosDelsy Dyane CtPas encore d'évaluation
- InmunoDocument8 pagesInmunoAndres ErasoPas encore d'évaluation
- INMUNOLOGIADocument4 pagesINMUNOLOGIAAnderson ValenciaPas encore d'évaluation
- Influencia de La MicrobiotaDocument13 pagesInfluencia de La MicrobiotaMilagros Elaine Blas PizarroPas encore d'évaluation
- Guerra Infinita - Trichomonas Vaginalis e Interacciones Con La Respuesta Inmune Del Huésped - PMC 1 ESPAÑOLDocument27 pagesGuerra Infinita - Trichomonas Vaginalis e Interacciones Con La Respuesta Inmune Del Huésped - PMC 1 ESPAÑOLMónica MartínezPas encore d'évaluation
- Un Comentario Acerca de Los Motivos CPGDocument5 pagesUn Comentario Acerca de Los Motivos CPGajdf5690Pas encore d'évaluation
- Articulo OMEGA 3Document8 pagesArticulo OMEGA 3Brenda ValdespinoPas encore d'évaluation
- 36.03 Fisiopatologia de La Rinitis AlérgicaDocument7 pages36.03 Fisiopatologia de La Rinitis AlérgicaCarlos Miguel GarzotaPas encore d'évaluation
- ResumenDocument118 pagesResumenKaren Ospina SalazarPas encore d'évaluation
- Fisiopatología de La SarcoidosisDocument12 pagesFisiopatología de La SarcoidosisClaudioMolinaPas encore d'évaluation
- Inmunidad Natural o InnataDocument2 pagesInmunidad Natural o InnataOlga Esther Salazar OsunaPas encore d'évaluation
- Caso Clínico 3 Respuesta InmuneDocument4 pagesCaso Clínico 3 Respuesta InmuneGiohanna Hervias BallonPas encore d'évaluation
- Mediadores Inflamatorios PDFDocument11 pagesMediadores Inflamatorios PDFkattializzetPas encore d'évaluation
- Phagocytosis of Microbes ESDocument53 pagesPhagocytosis of Microbes ESStephani LopezPas encore d'évaluation
- Cuestionario Respuesta InmunitariaDocument5 pagesCuestionario Respuesta InmunitariaMercedes CamPas encore d'évaluation
- E. ColiDocument5 pagesE. ColiDaniela GonzálezPas encore d'évaluation
- 2022petakn Et Al Weil's Disease, Leptospirosis EsDocument27 pages2022petakn Et Al Weil's Disease, Leptospirosis EsEdwar ÑustesPas encore d'évaluation
- Homeostasis Intestinal - Colaboración Del Sistema Inmune Con La MicrobiotaDocument5 pagesHomeostasis Intestinal - Colaboración Del Sistema Inmune Con La MicrobiotaIVAN JACOB RAMOS REYESPas encore d'évaluation
- Transferencia de inmunidad en el binomio madre-cría: Estudio comparativo entre especiesD'EverandTransferencia de inmunidad en el binomio madre-cría: Estudio comparativo entre especiesPas encore d'évaluation
- AmiloidosisDocument5 pagesAmiloidosisRouz TorresPas encore d'évaluation
- CorticoDocument4 pagesCorticoRouz TorresPas encore d'évaluation
- FLUOROQUINOLONASDocument2 pagesFLUOROQUINOLONASRouz TorresPas encore d'évaluation
- Antibioticos MacrolidosDocument5 pagesAntibioticos MacrolidosRouz TorresPas encore d'évaluation
- DiureticosDocument3 pagesDiureticosRouz TorresPas encore d'évaluation
- AntihipertensivosDocument5 pagesAntihipertensivosRouz TorresPas encore d'évaluation
- Aines2 0Document8 pagesAines2 0Rouz TorresPas encore d'évaluation
- FormasfarmaceuticasDocument1 pageFormasfarmaceuticasRouz TorresPas encore d'évaluation
- 3 GeneraciónDocument2 pages3 GeneraciónRouz TorresPas encore d'évaluation
- Antibioticos PenicilinasDocument2 pagesAntibioticos PenicilinasRouz TorresPas encore d'évaluation
- DERMATOLOGÍADocument6 pagesDERMATOLOGÍARouz TorresPas encore d'évaluation
- Daños A La Salud de La Población Del ÁreaDocument14 pagesDaños A La Salud de La Población Del ÁreaRouz TorresPas encore d'évaluation
- Fiebre TifoideaDocument1 pageFiebre TifoideaRouz TorresPas encore d'évaluation
- Biodegradacion 130808163348 Phpapp02Document13 pagesBiodegradacion 130808163348 Phpapp02Rouz TorresPas encore d'évaluation
- Municipio de Juan GalindoDocument3 pagesMunicipio de Juan GalindoRouz TorresPas encore d'évaluation
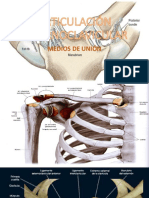
- Articulación EsternoclavicularDocument10 pagesArticulación EsternoclavicularRouz TorresPas encore d'évaluation
- Movimiento AntiglobalizaciónDocument4 pagesMovimiento AntiglobalizaciónRouz TorresPas encore d'évaluation
- Citometria HemáticaDocument17 pagesCitometria HemáticaRouz TorresPas encore d'évaluation
- Aines2 0Document8 pagesAines2 0Rouz TorresPas encore d'évaluation
- Movimiento FeministaDocument3 pagesMovimiento FeministaRouz TorresPas encore d'évaluation
- Tareas de La Unidad IIDocument43 pagesTareas de La Unidad IIRouz TorresPas encore d'évaluation
- Trastornos de AnsiedadDocument19 pagesTrastornos de AnsiedadRouz TorresPas encore d'évaluation
- Guía de Estudios PAA para América LatinaDocument75 pagesGuía de Estudios PAA para América LatinaAndrès FloresPas encore d'évaluation
- Arterias Del EncéfaloDocument5 pagesArterias Del EncéfaloRouz TorresPas encore d'évaluation
- BrucellaDocument13 pagesBrucellaRouz TorresPas encore d'évaluation
- Articulación EsternoclavicularDocument10 pagesArticulación EsternoclavicularRouz TorresPas encore d'évaluation
- Casino RoyaleDocument38 pagesCasino RoyaleRouz TorresPas encore d'évaluation
- Iv. MacromolculasDocument44 pagesIv. MacromolculasDaniel FernándezPas encore d'évaluation
- Paranthropus RobustusDocument19 pagesParanthropus RobustusNicolas Patricio SitjaPas encore d'évaluation
- Con Tam in AntesDocument23 pagesCon Tam in AnteslucasynicolasPas encore d'évaluation
- Virus de La Varicela ZósterDocument46 pagesVirus de La Varicela ZósterEdwin Ramiro Torrico VargasPas encore d'évaluation
- GUIA DIDACTICA No 5 El CerebroDocument15 pagesGUIA DIDACTICA No 5 El Cerebrooglb1970Pas encore d'évaluation
- Perros TripticoDocument2 pagesPerros TripticoRosita Maria Mercedes100% (2)
- Cemento Efectos en La SaludDocument31 pagesCemento Efectos en La SaludLuis Pintado Sanchez100% (1)
- Competencias 16 Agosto 2013 Definitivo 2Document53 pagesCompetencias 16 Agosto 2013 Definitivo 2YENNY ALEJANDRA NEGRETE SANCHEZPas encore d'évaluation
- Ensayo Experimentación AnimalDocument4 pagesEnsayo Experimentación AnimalAngelica Triana0% (1)
- Los DelfinesDocument1 pageLos DelfinesArnaldoloPas encore d'évaluation
- Trabajo Sist CirculatorioDocument5 pagesTrabajo Sist CirculatorioJose Luis Salas Cia100% (1)
- Paseos Escolares 2018Document12 pagesPaseos Escolares 2018Guido MoisesPas encore d'évaluation
- Guia Ciencias Naturales Grado 2° Semana 1 Al 5 MarisolDocument11 pagesGuia Ciencias Naturales Grado 2° Semana 1 Al 5 MarisolDevis germán Beltrán JiménezPas encore d'évaluation
- Triangulos Del CuelloDocument9 pagesTriangulos Del CuelloErick Montenegro100% (1)
- Preguntas ENAMDocument3 pagesPreguntas ENAMMilagros Montenegro CruzPas encore d'évaluation
- ImagenDocument113 pagesImagenLeslie Yuz AracenaPas encore d'évaluation
- Informe de Practica de Produccion y Salud de Ovinos (Parasito)Document13 pagesInforme de Practica de Produccion y Salud de Ovinos (Parasito)Jorge Janampa CamposPas encore d'évaluation
- Sistema Circulatorio de Los Insectos.Document4 pagesSistema Circulatorio de Los Insectos.Carlos Alberto Hernández TorresPas encore d'évaluation
- Testimonio Padre ChorrDocument4 pagesTestimonio Padre Chorrapi-3699819100% (1)
- Resultado PARÁSITOSDocument2 pagesResultado PARÁSITOSKenny Grau RodriguezPas encore d'évaluation
- Trauma DentalDocument32 pagesTrauma DentalGabriel García100% (3)
- SilaboDocument13 pagesSilaboJuanaPas encore d'évaluation
- Los Discipulos de Cthulhu - A A AttanasioDocument116 pagesLos Discipulos de Cthulhu - A A AttanasioAnonymous 6pkqSSm100% (2)
- Dossier Curso Adiestrador Canino 22-23Document20 pagesDossier Curso Adiestrador Canino 22-23Thais LaespadaPas encore d'évaluation
- LECTURA CRITICA SéptimoDocument5 pagesLECTURA CRITICA SéptimoJhon AgudeloPas encore d'évaluation
- Ca OvarioDocument84 pagesCa OvarioFranck PalacinPas encore d'évaluation
- Plano CAMAL 18 08 2017 ALPACAS PDFDocument1 pagePlano CAMAL 18 08 2017 ALPACAS PDFgiomar leon gutierrezPas encore d'évaluation
- Preguntas GastroenterologiaDocument5 pagesPreguntas GastroenterologiaDaniel CoriaPas encore d'évaluation
- Informe de Helado de Frutas Grupo 2Document8 pagesInforme de Helado de Frutas Grupo 2suyiPas encore d'évaluation
- 4to Fisiologia RespiratoriaDocument1 page4to Fisiologia RespiratoriaAna CastillaPas encore d'évaluation
- Tracción Humana y Las Herramientas Que UtilizaDocument16 pagesTracción Humana y Las Herramientas Que UtilizaJORGE EDUARDO JIMENEZ CRUZ50% (2)