Vous aimerez peut-être aussi
- Basic Data EntryDocument12 pagesBasic Data EntryVimal Anbalagan0% (1)
- Lecture 1-2 Vacuum SystemDocument113 pagesLecture 1-2 Vacuum SystemMehreen AkmalPas encore d'évaluation
- Sucrose - Properties and ApplicationsDocument307 pagesSucrose - Properties and ApplicationsSharini_Mu_oz_137100% (1)
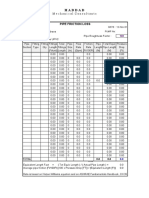
- Custom Engineered Piping Data ReportDocument94 pagesCustom Engineered Piping Data ReportshaonaaPas encore d'évaluation
- Calculate Gas Compressibility Excel FunctionDocument2 pagesCalculate Gas Compressibility Excel FunctionCHANADASPas encore d'évaluation
- Piping PDSDocument2 pagesPiping PDSsunii19847908Pas encore d'évaluation
- HKDSE Listening Part B TechniquesDocument40 pagesHKDSE Listening Part B TechniquesJayantPas encore d'évaluation
- Thermal Properties of Matter: Internal Energy, Heat Capacity, Phase ChangesDocument79 pagesThermal Properties of Matter: Internal Energy, Heat Capacity, Phase ChangesHafiza JaweriaPas encore d'évaluation
- How To Use The Excel "Linest" Function For Linear Regression Models With Multiple Input Variables XDocument8 pagesHow To Use The Excel "Linest" Function For Linear Regression Models With Multiple Input Variables XANIL PALPas encore d'évaluation
- Viscosity (SI)Document6 pagesViscosity (SI)Liem Handi Wijaya SusantoPas encore d'évaluation
- Unit Conversion All in OneDocument32 767 pagesUnit Conversion All in OneAnonymous dRl45itPas encore d'évaluation
- CSB DetailsDocument52 pagesCSB DetailsVelmurugan ElumalaiPas encore d'évaluation
- Conversion Factors: A Comprehensive Guide to Measurement UnitsDocument11 pagesConversion Factors: A Comprehensive Guide to Measurement Unitsmicro kidPas encore d'évaluation
- Yaws - Chemical Properties Handbook Physical, Thermodynamics, Engironmental Transport, Safety & Health Related Properties For Organic & Inorganic ChemicalDocument2 pagesYaws - Chemical Properties Handbook Physical, Thermodynamics, Engironmental Transport, Safety & Health Related Properties For Organic & Inorganic ChemicalRanny NovellaPas encore d'évaluation
- PEBD-EVA Blends PDFDocument9 pagesPEBD-EVA Blends PDFDiana Carolina CalvoPas encore d'évaluation
- Density Viscosity and Surface Tension of Five Vegetable Oils at Elevated Temperatures Measurement and Modeling PDFDocument18 pagesDensity Viscosity and Surface Tension of Five Vegetable Oils at Elevated Temperatures Measurement and Modeling PDFSnigdha SarmahPas encore d'évaluation
- Excel PolynomialDocument5 pagesExcel Polynomial______.________Pas encore d'évaluation
- US Bone Gelatin Sourcing and ManufacturingDocument26 pagesUS Bone Gelatin Sourcing and ManufacturingTapan SharmaPas encore d'évaluation
- Enthalpy / Entropy DiagramDocument34 pagesEnthalpy / Entropy DiagramMara TrofinPas encore d'évaluation
- Piping Specialty Worksheets IndexDocument110 pagesPiping Specialty Worksheets IndexmanhngoducPas encore d'évaluation
- Determining Specific Heat Capacity Using a Heating CoilDocument32 pagesDetermining Specific Heat Capacity Using a Heating CoilMartinPas encore d'évaluation
- Solid Bulk DensityDocument21 pagesSolid Bulk DensityMauricio RodriguezPas encore d'évaluation
- Apendice 2Document12 pagesApendice 2JA UMAN1Pas encore d'évaluation
- Oyster Mushroom PolvoronDocument14 pagesOyster Mushroom PolvoronMark Lloyd Argis100% (3)
- Milk ProductsDocument16 pagesMilk ProductsTeodora AncaPas encore d'évaluation
- Review: Gelatin, Source, Extraction and Industrial ApplicationsDocument14 pagesReview: Gelatin, Source, Extraction and Industrial ApplicationsJade NguyenPas encore d'évaluation
- Library WebDocument128 pagesLibrary WebVikash AgarwalPas encore d'évaluation
- Freeze Protection: Heat TracingDocument4 pagesFreeze Protection: Heat TracingNaser JahangiriPas encore d'évaluation
- Water - Dynamic and Kinematic ViscosityDocument3 pagesWater - Dynamic and Kinematic ViscosityManik KandanPas encore d'évaluation
- Raz Lq40 Goliathbirdeatingspidersgiantarachnids CLRDocument9 pagesRaz Lq40 Goliathbirdeatingspidersgiantarachnids CLRaprilia rahmawatiPas encore d'évaluation
- Thermal Properties of FoodsDocument12 pagesThermal Properties of FoodspedroloxxxPas encore d'évaluation
- Thermal Properties of FoodsDocument12 pagesThermal Properties of FoodspedroloxxxPas encore d'évaluation
- Appendix D-2 Viscosity of Pure Water 0-100° PDFDocument1 pageAppendix D-2 Viscosity of Pure Water 0-100° PDFLuis Argüelles SaenzPas encore d'évaluation
- Yaws Chapter 1Document1 pageYaws Chapter 1basana siamremarePas encore d'évaluation
- TLE 10 Q1 Lesson 1Document23 pagesTLE 10 Q1 Lesson 1Angustia HalogPas encore d'évaluation
- How To Estimate The Heat Penetration Factor FH of Packaged Foods PDFDocument71 pagesHow To Estimate The Heat Penetration Factor FH of Packaged Foods PDFAlex Samuel SilvaPas encore d'évaluation
- Latent Heat of Vaporization of Ethanol Using Clausius-Clapeyron EquationDocument5 pagesLatent Heat of Vaporization of Ethanol Using Clausius-Clapeyron EquationMel DyPas encore d'évaluation
- Caking of Raw SugarDocument52 pagesCaking of Raw Sugarmarcelo22061970Pas encore d'évaluation
- WayToEnglish4 - ANK - SB - 19747 2Document31 pagesWayToEnglish4 - ANK - SB - 19747 2DA MAPas encore d'évaluation
- 2010 - Thermal Process Calculations Through Ball's Original FormulaDocument16 pages2010 - Thermal Process Calculations Through Ball's Original FormulapedroloxxxPas encore d'évaluation
- Moisture Dependent Thermal Properties of Doum PalmDocument6 pagesMoisture Dependent Thermal Properties of Doum Palmgaza manPas encore d'évaluation
- Data fitting in Scilab: interpolation and approximation techniquesDocument16 pagesData fitting in Scilab: interpolation and approximation techniquesAlexander Diaz AlvarezPas encore d'évaluation
- Moisture Content of Natural GasesDocument8 pagesMoisture Content of Natural Gasesarafat latifPas encore d'évaluation
- Chapter 20 The First Law of ThermodynamicsDocument78 pagesChapter 20 The First Law of ThermodynamicsNeil de DiosPas encore d'évaluation
- S SDocument41 pagesS SKristi RogersPas encore d'évaluation
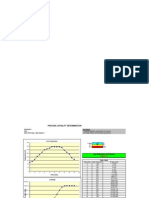
- Process Lethality Spreadsheet - 2Document8 pagesProcess Lethality Spreadsheet - 2TranVanHoaPas encore d'évaluation
- Process Lethality Determination Example:: Salmonella Salmonella E. Coli O157:H7 Lean Gr. Beef (2% Fat)Document1 pageProcess Lethality Determination Example:: Salmonella Salmonella E. Coli O157:H7 Lean Gr. Beef (2% Fat)Mr ThanhPas encore d'évaluation
- Introduction To Food Freezing - 3Document50 pagesIntroduction To Food Freezing - 3tadsrikanth_chainlu100% (1)
- Meat ProductsDocument18 pagesMeat ProductsChristian Oribe CastilloPas encore d'évaluation
- LR-Practical 8 (Texture)Document12 pagesLR-Practical 8 (Texture)najwaPas encore d'évaluation
- Removing fat from meat plant wastewater with hydrocyclonesDocument63 pagesRemoving fat from meat plant wastewater with hydrocyclonesNopparutPas encore d'évaluation
- Visualizing Non-Newtonian Fluid Properties with OobleckDocument6 pagesVisualizing Non-Newtonian Fluid Properties with OobleckAditya.H.MPas encore d'évaluation
- Chapter 2 - History of Dimensional AnalysisDocument7 pagesChapter 2 - History of Dimensional AnalysisShaikh Abdur RazzakPas encore d'évaluation
- 1 Artigo Heat and Mass Transfer in Industrial Biscuit Baking Oven and Effect ofDocument12 pages1 Artigo Heat and Mass Transfer in Industrial Biscuit Baking Oven and Effect ofMarianne Soares100% (1)
- Droplet Evaporation With Reference To The Effectiveness of Water Mist CoolingDocument9 pagesDroplet Evaporation With Reference To The Effectiveness of Water Mist CoolingWhidas PrihantoroPas encore d'évaluation
- Drying Characteristics of Mango Slices Using The Refractance Window™ Technique PDFDocument7 pagesDrying Characteristics of Mango Slices Using The Refractance Window™ Technique PDFcedu126Pas encore d'évaluation
- Dehydration of MilkDocument23 pagesDehydration of MilkMohammad Ashraf Paul50% (2)
- Adsorption Column Specs Rev1Document6 pagesAdsorption Column Specs Rev1dedyPas encore d'évaluation
- ChE Industrial Chem Manual Final - 002Document37 pagesChE Industrial Chem Manual Final - 002rossangel35Pas encore d'évaluation
- Computational Analysis of Thermal Behavior Within A Scraped Surface Heat Exchanger SSHEDocument9 pagesComputational Analysis of Thermal Behavior Within A Scraped Surface Heat Exchanger SSHEEditor IJTSRDPas encore d'évaluation
- Filtration Unit Report: Index Terms-Pressure, Flow, Pump, Filtration, ParticleDocument4 pagesFiltration Unit Report: Index Terms-Pressure, Flow, Pump, Filtration, Particlejuan francoPas encore d'évaluation
- Hs DiagramDocument32 pagesHs DiagramsubratorajPas encore d'évaluation
- Materials of ConstructionDocument3 pagesMaterials of ConstructionMary Antonette M. TingalPas encore d'évaluation
- Determination of Ash in BiomassDocument8 pagesDetermination of Ash in Biomassmarta_trninicPas encore d'évaluation
- Recommended Laboratory Analytical Procedures For DDGSDocument4 pagesRecommended Laboratory Analytical Procedures For DDGSAulia VellaraniPas encore d'évaluation
- Supercooling Preservation On Quality of Beef SteakDocument39 pagesSupercooling Preservation On Quality of Beef SteakThuy LePas encore d'évaluation
- JURIN - Effect of Novel Quick Freezing Techniques CombinedDocument7 pagesJURIN - Effect of Novel Quick Freezing Techniques CombinedWahyu Trimurti HandayaniPas encore d'évaluation
- Masa Simpan 18 BulanDocument10 pagesMasa Simpan 18 BulanRiri JuniartiPas encore d'évaluation
- Metal Alloy Used For Food ContactDocument218 pagesMetal Alloy Used For Food ContactLukePas encore d'évaluation
- NZ MPI Vibrio Vulnificus - Microbial Pathogen Data SheetDocument3 pagesNZ MPI Vibrio Vulnificus - Microbial Pathogen Data SheetpedroloxxxPas encore d'évaluation
- SporeDocument8 pagesSporepedroloxxxPas encore d'évaluation
- AppertDocument158 pagesAppertpedroloxxxPas encore d'évaluation
- Eurobarometer 2010 - Report PDFDocument168 pagesEurobarometer 2010 - Report PDFpedroloxxxPas encore d'évaluation
- 2017 - Thermal Properties of Skipjack TunaDocument12 pages2017 - Thermal Properties of Skipjack TunapedroloxxxPas encore d'évaluation
- NSSP Guide for Safe ShellfishDocument516 pagesNSSP Guide for Safe ShellfishpedroloxxxPas encore d'évaluation
- Clostridium Botulinum, Clostridia: and NeurotoxigenicDocument4 pagesClostridium Botulinum, Clostridia: and NeurotoxigenicpedroloxxxPas encore d'évaluation
- 2017 - Thermal Properties of Skipjack TunaDocument7 pages2017 - Thermal Properties of Skipjack TunapedroloxxxPas encore d'évaluation
- 20xx - Ball's Formula Method RevisitedDocument6 pages20xx - Ball's Formula Method RevisitedpedroloxxxPas encore d'évaluation
- 20xx - Ball's Formula Method RevisitedDocument6 pages20xx - Ball's Formula Method RevisitedpedroloxxxPas encore d'évaluation
- HACCP BookletDocument30 pagesHACCP BookletMaria DapkeviciusPas encore d'évaluation
- EFSA - Risk Assessment Vs Risk Management PDFDocument1 pageEFSA - Risk Assessment Vs Risk Management PDFpedroloxxxPas encore d'évaluation
- FDA - Potentially Hazardous Foods PDFDocument3 pagesFDA - Potentially Hazardous Foods PDFpedroloxxxPas encore d'évaluation
- FDA - Potentially Hazardous Foods PDFDocument3 pagesFDA - Potentially Hazardous Foods PDFpedroloxxxPas encore d'évaluation
- Big Bug BookDocument292 pagesBig Bug BookAnjukaPas encore d'évaluation
- FDA - Potentially Hazardous FoodsDocument4 pagesFDA - Potentially Hazardous FoodspedroloxxxPas encore d'évaluation
- 2015 - The Check Problem of Food Thermal Processes. A Mathematical SolutionDocument29 pages2015 - The Check Problem of Food Thermal Processes. A Mathematical SolutionpedroloxxxPas encore d'évaluation
- 2015 - The Check Problem of Food Thermal Processes. A Mathematical SolutionDocument15 pages2015 - The Check Problem of Food Thermal Processes. A Mathematical SolutionpedroloxxxPas encore d'évaluation
- 2015 - The Check Problem of Food Thermal Processes. A Mathematical SolutionDocument15 pages2015 - The Check Problem of Food Thermal Processes. A Mathematical SolutionpedroloxxxPas encore d'évaluation
- Review Histamine ProducingbacteriaDocument10 pagesReview Histamine ProducingbacteriapedroloxxxPas encore d'évaluation
- 2017 - Thermal Properties of Skipjack TunaDocument7 pages2017 - Thermal Properties of Skipjack TunapedroloxxxPas encore d'évaluation
- 2010 - Retort Cooling Water Bacteriological LoadDocument8 pages2010 - Retort Cooling Water Bacteriological LoadpedroloxxxPas encore d'évaluation
- 2013 - A New Mathematical Model For Food Thermal Process PredictionDocument9 pages2013 - A New Mathematical Model For Food Thermal Process PredictionpedroloxxxPas encore d'évaluation
- 2012 - Evaluación y Optimizacion de Ltratamiento TermicoDocument17 pages2012 - Evaluación y Optimizacion de Ltratamiento TermicopedroloxxxPas encore d'évaluation
- 2010 - Retort Cooling Water Bacteriological LoadDocument8 pages2010 - Retort Cooling Water Bacteriological LoadpedroloxxxPas encore d'évaluation
- Kazan Caffe Bar price listDocument6 pagesKazan Caffe Bar price listIvanaPas encore d'évaluation
- Livelihood Promotion: Sectors, Interventions & StrategiesDocument31 pagesLivelihood Promotion: Sectors, Interventions & StrategiesSri Himaja100% (1)
- FPC Fresh Awards Guest List Programme FINAL 15.09.2022Document17 pagesFPC Fresh Awards Guest List Programme FINAL 15.09.2022m2Pas encore d'évaluation
- Event Checklist FormDocument10 pagesEvent Checklist FormJonathan D'OliveiroPas encore d'évaluation
- TraditionsDocument3 pagesTraditionssherina vinoyaPas encore d'évaluation
- The Bond of Love - Kenneth AndersonDocument7 pagesThe Bond of Love - Kenneth Andersonsnehaa sPas encore d'évaluation
- Oum-Bbek1103 (Principles of Microeconomics)Document21 pagesOum-Bbek1103 (Principles of Microeconomics)jesson kimPas encore d'évaluation
- Chapter 101-160Document297 pagesChapter 101-160Dipankar BoruahPas encore d'évaluation
- Freeze-dried food and urban wildlifeDocument6 pagesFreeze-dried food and urban wildlifedanialPas encore d'évaluation
- B A-B Sc-SyllabusDocument26 pagesB A-B Sc-SyllabusAkshat SharmaPas encore d'évaluation
- A 1000 Calorie Diet Plan For Weight Loss - DMooseDocument15 pagesA 1000 Calorie Diet Plan For Weight Loss - DMoosekwasnyciakristinaPas encore d'évaluation
- Passive Voice Simple Present Tense WorksheetDocument2 pagesPassive Voice Simple Present Tense WorksheetSegundo AlmagroPas encore d'évaluation
- BasterDocument10 pagesBasterAimae Eata EalaPas encore d'évaluation
- CPC Yao Dao Primary School P.5 Grammar Notes: Name: - ClassDocument25 pagesCPC Yao Dao Primary School P.5 Grammar Notes: Name: - ClassTracy ChowPas encore d'évaluation
- Shuttle Routes and DriversDocument23 pagesShuttle Routes and DriversJazz AmiryPas encore d'évaluation
- Mapeh 8 4Q Final ExamDocument5 pagesMapeh 8 4Q Final ExamJennifer DagatanPas encore d'évaluation
- Filipino Rice Farmers Face Poverty Despite Vital RoleDocument3 pagesFilipino Rice Farmers Face Poverty Despite Vital RoleMike JohnPas encore d'évaluation
- Tom Kropp "Bad Betty"Document3 pagesTom Kropp "Bad Betty"Barbara BergmannPas encore d'évaluation
- CH 10 AshtangaDocument12 pagesCH 10 AshtangarushitPas encore d'évaluation
- Rituparna Banerjee, Arun K. Verma, Mohammed Wasim Siddiqui - Natural Antioxidants - Applications in Foods of Animal Origin-Apple Academic Press (2017)Document414 pagesRituparna Banerjee, Arun K. Verma, Mohammed Wasim Siddiqui - Natural Antioxidants - Applications in Foods of Animal Origin-Apple Academic Press (2017)wilmerjmorenoPas encore d'évaluation
- And Everyone Wins (Bechloe)Document276 pagesAnd Everyone Wins (Bechloe)mercedesmgPas encore d'évaluation
- Bahasa Inggris Untuk Bahasa Indonesia PDFDocument85 pagesBahasa Inggris Untuk Bahasa Indonesia PDFdamarPas encore d'évaluation
- Robinson Crusoe in Levels Elementary LevelDocument67 pagesRobinson Crusoe in Levels Elementary LevelWardaPas encore d'évaluation
- RC Cola Marketing Plan DocumentationDocument28 pagesRC Cola Marketing Plan DocumentationRhian Lacoste77% (13)
- How The Monkey Got A Drink (Brazilian Fariy Tale)Document8 pagesHow The Monkey Got A Drink (Brazilian Fariy Tale)Kacper KruszewskiPas encore d'évaluation