Vous aimerez peut-être aussi
- Defining Physiology: Principles, Themes, Concepts.: Hwee Ming Cheng Kin Kheong Mah Kumar SeluakumaranDocument104 pagesDefining Physiology: Principles, Themes, Concepts.: Hwee Ming Cheng Kin Kheong Mah Kumar SeluakumaranGiovanni HenryPas encore d'évaluation
- Reirradiation Meta Analysis and Systematic ReviewDocument13 pagesReirradiation Meta Analysis and Systematic ReviewGiovanni HenryPas encore d'évaluation
- Tracheal StenosisDocument5 pagesTracheal StenosisGiovanni HenryPas encore d'évaluation
- Anterior Pharyngeal PouchDocument4 pagesAnterior Pharyngeal PouchGiovanni HenryPas encore d'évaluation
- Paediatric Neck MassesDocument7 pagesPaediatric Neck MassesGiovanni HenryPas encore d'évaluation
- 2018 Cardiovascular PhysiologyDocument197 pages2018 Cardiovascular PhysiologyGladys ParedesPas encore d'évaluation
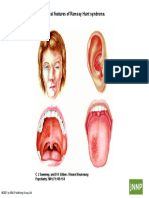
- Clinical Features of Ramsay Hunt Syndrome.: C J Sweeney, and D H Gilden J Neurol Neurosurg Psychiatry 2001 71:149-154Document1 pageClinical Features of Ramsay Hunt Syndrome.: C J Sweeney, and D H Gilden J Neurol Neurosurg Psychiatry 2001 71:149-154Giovanni HenryPas encore d'évaluation
- Evaluation of The Dizzy PatientDocument61 pagesEvaluation of The Dizzy PatientGiovanni HenryPas encore d'évaluation
- Endoscopic Sinus SurgeryDocument91 pagesEndoscopic Sinus SurgeryGiovanni HenryPas encore d'évaluation
- April 2018 Recalls - Reda: Embolectomy??)Document5 pagesApril 2018 Recalls - Reda: Embolectomy??)Giovanni HenryPas encore d'évaluation
- Defining Physiology: Principles, Themes, Concepts.: Hwee Ming Cheng Kin Kheong Mah Kumar SeluakumaranDocument104 pagesDefining Physiology: Principles, Themes, Concepts.: Hwee Ming Cheng Kin Kheong Mah Kumar SeluakumaranGiovanni HenryPas encore d'évaluation
- Sept 2020 MRCS RecallsDocument46 pagesSept 2020 MRCS RecallsGiovanni Henry75% (4)
- 6th Central Pay Commission Salary CalculatorDocument15 pages6th Central Pay Commission Salary Calculatorrakhonde100% (436)
- Recalls of Paper 2 - 11/9/2018: ConsentDocument6 pagesRecalls of Paper 2 - 11/9/2018: ConsentGiovanni Henry0% (1)
- September 2020 Recalls Scorpio and Al Beruni 243 QuestionsDocument9 pagesSeptember 2020 Recalls Scorpio and Al Beruni 243 QuestionsGiovanni HenryPas encore d'évaluation
- April 2018 IMRCS A Exam ReviewDocument12 pagesApril 2018 IMRCS A Exam Reviewmobz00Pas encore d'évaluation
- Medical EMQs and Clinical ScenariosDocument18 pagesMedical EMQs and Clinical ScenariosMehdi Hasan MazumderPas encore d'évaluation
- The Physiology of Human DefecationDocument21 pagesThe Physiology of Human Defecationyohanna yohannaPas encore d'évaluation
- Transplantation: Leo-Paul Powell DM Tutorial Series March 30, 2021Document45 pagesTransplantation: Leo-Paul Powell DM Tutorial Series March 30, 2021Giovanni HenryPas encore d'évaluation
- DNB Gen Surgery Old Question Papers 1Document28 pagesDNB Gen Surgery Old Question Papers 1sidorai100% (1)
- Surgical AnatomyDocument4 pagesSurgical AnatomyGiovanni HenryPas encore d'évaluation
- Endothelium I: Atherogenesis: Maintenance of Vascular HomeostasisDocument5 pagesEndothelium I: Atherogenesis: Maintenance of Vascular HomeostasisGiovanni HenryPas encore d'évaluation
- Gen Surg 1996 2009Document26 pagesGen Surg 1996 2009LijoeliyasPas encore d'évaluation
- General Surgery June 2010: Paper IDocument2 pagesGeneral Surgery June 2010: Paper IGiovanni HenryPas encore d'évaluation
- Surgery Questions EssayDocument11 pagesSurgery Questions Essaymimycutey100% (3)
- EVAR vs. Open Repair - EvidenceDocument5 pagesEVAR vs. Open Repair - EvidenceGiovanni HenryPas encore d'évaluation
- DNB Gen Surgery Old Question Papers 1Document28 pagesDNB Gen Surgery Old Question Papers 1sidorai100% (1)
- Colon & Rectum Questions Aug 2008Document61 pagesColon & Rectum Questions Aug 2008Dr.mesho Bin MansourPas encore d'évaluation
- Class of 2023 Year 1 Sem 1 Final Aug 30Document16 pagesClass of 2023 Year 1 Sem 1 Final Aug 30Giovanni HenryPas encore d'évaluation
- Frcs Viva TOPICSDocument4 pagesFrcs Viva TOPICSGiovanni HenryPas encore d'évaluation
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (894)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (265)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (119)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Autonomic Nervous System: Prof. DRDocument65 pagesAutonomic Nervous System: Prof. DRGo HellPas encore d'évaluation
- Group 10: Peripheral Nervous System (PNS)Document24 pagesGroup 10: Peripheral Nervous System (PNS)Tara DolotaPas encore d'évaluation
- Flashcards - Lower LimbDocument6 pagesFlashcards - Lower LimbKhanPas encore d'évaluation
- PG Syllabus 2nd Year PDFDocument95 pagesPG Syllabus 2nd Year PDFDevrajPas encore d'évaluation
- Nervous System Answers to 3.10 QuestionsDocument1 pageNervous System Answers to 3.10 QuestionsbrownieallennPas encore d'évaluation
- Sensory and Motor Cranial NervesDocument13 pagesSensory and Motor Cranial NervesAnn HeerahPas encore d'évaluation
- Michael Shacklock, M.app - SC, Dip. Physio. (Auth.) - Clinical Neurodynamics. A New System of Musculoskeletal TreatmentDocument255 pagesMichael Shacklock, M.app - SC, Dip. Physio. (Auth.) - Clinical Neurodynamics. A New System of Musculoskeletal TreatmentІгор Сомов100% (1)
- Johanes Hardjono: Problematik Fisioterapi Dan Diagnose FisioterapiDocument30 pagesJohanes Hardjono: Problematik Fisioterapi Dan Diagnose FisioterapisulaimanPas encore d'évaluation
- (Douglas W. Zochodne) Neurobiology of Peripheral NDocument318 pages(Douglas W. Zochodne) Neurobiology of Peripheral NImmer Armendáriz BetancourtPas encore d'évaluation
- Peripheral NeuropathyDocument8 pagesPeripheral NeuropathyJessica Alejandra Razo100% (1)
- Organization of NSDocument20 pagesOrganization of NSnadine azmyPas encore d'évaluation
- Trigeminal NerveDocument25 pagesTrigeminal Nerveبراءة أحمد السلاماتPas encore d'évaluation
- The Nervous System and Reflex Arc (GCSE)Document35 pagesThe Nervous System and Reflex Arc (GCSE)Raja UsamaPas encore d'évaluation
- Mandibular NerveDocument6 pagesMandibular NerveCatalina StoicescuPas encore d'évaluation
- Science Form 3 CHAPTER 1Document8 pagesScience Form 3 CHAPTER 1SOLEHAH BINTI MAT DAUD Moe85% (13)
- @anesthesia - Books 2015 Basic Physiology For Anaesthetists PDFDocument455 pages@anesthesia - Books 2015 Basic Physiology For Anaesthetists PDFovi67% (3)
- Introduction To Nerve Conduction StudiesDocument15 pagesIntroduction To Nerve Conduction StudiesIngrid GomesPas encore d'évaluation
- Q2 Week 4 Day 2Document4 pagesQ2 Week 4 Day 2chona redillasPas encore d'évaluation
- A Short Guide to Understanding FibromyalgiaDocument188 pagesA Short Guide to Understanding FibromyalgiaDave100% (4)
- Peripheral Nerve Injuries ExplainedDocument4 pagesPeripheral Nerve Injuries ExplainedJianni ClaridadPas encore d'évaluation
- 111Document80 pages111Amrut HosmaniPas encore d'évaluation
- Basic Brain AnatomyDocument36 pagesBasic Brain Anatomymartha wibawaPas encore d'évaluation
- Understanding the Complex Nervous SystemDocument43 pagesUnderstanding the Complex Nervous Systemwieka mawiePas encore d'évaluation
- Activity 1: Break It Down Activity 3: The Nerves!!!Document2 pagesActivity 1: Break It Down Activity 3: The Nerves!!!Jia CapatiPas encore d'évaluation
- Conflicting Dermatome Maps Clinical ImplicationsDocument8 pagesConflicting Dermatome Maps Clinical ImplicationsPedro BaptistaPas encore d'évaluation
- Spinal Disorders - Study GuideDocument183 pagesSpinal Disorders - Study Guidetechsupport24712365Pas encore d'évaluation
- Cranial Nerve V2 - Maxillary Nerve - FRCEM SuccessDocument1 pageCranial Nerve V2 - Maxillary Nerve - FRCEM SuccessJohn CoxPas encore d'évaluation
- Neuro IDocument6 pagesNeuro IElenaPas encore d'évaluation
- Entrapment Neuropathy Entrapment NeuropathyDocument36 pagesEntrapment Neuropathy Entrapment NeuropathyNEuRoLoGisT CoFFeeCuP100% (1)
- Dietrich Klighardt & Daniela Deiosso - A.R.T. 1 Beginners ManualDocument74 pagesDietrich Klighardt & Daniela Deiosso - A.R.T. 1 Beginners Manualbjjman100% (3)