Vous aimerez peut-être aussi
- Carbohydrate Chemistry—8: Plenary Lectures Presented at the Eighth International Symposium on Carbohydrate Chemistry, Kyoto, Japan 16 - 20 August 1976D'EverandCarbohydrate Chemistry—8: Plenary Lectures Presented at the Eighth International Symposium on Carbohydrate Chemistry, Kyoto, Japan 16 - 20 August 1976K. OnoderaPas encore d'évaluation
- Chemesthesis: Chemical Touch in Food and EatingD'EverandChemesthesis: Chemical Touch in Food and EatingShane T. McDonaldPas encore d'évaluation
- Lipid Peroxidation Inhibition and Antiradical Activities of Some Leaf Fractions of Mangifera IndicaDocument7 pagesLipid Peroxidation Inhibition and Antiradical Activities of Some Leaf Fractions of Mangifera IndicaRoy WilsonPas encore d'évaluation
- Inhibition of Tetrahydrolipstatin: Pancreatic inDocument5 pagesInhibition of Tetrahydrolipstatin: Pancreatic inPatrick Del PradoPas encore d'évaluation
- Synthesis and Antimuscarinic Activity of 2-WEthyl-N-lhydroxyethylaminoethyl 22diphenylpropionate A Metaboliteof Aprophen jps.2600820603Document2 pagesSynthesis and Antimuscarinic Activity of 2-WEthyl-N-lhydroxyethylaminoethyl 22diphenylpropionate A Metaboliteof Aprophen jps.2600820603THEUSER0001Pas encore d'évaluation
- Food Chemistry: Hong Gao, Yi-Na Huang, Bo Gao, Pei-Yu Xu, Chika Inagaki, Jun KawabataDocument7 pagesFood Chemistry: Hong Gao, Yi-Na Huang, Bo Gao, Pei-Yu Xu, Chika Inagaki, Jun KawabataRayyhana ShopPas encore d'évaluation
- Stability and Enzymatic Studies With Omeprazole:Hydroxypropyl - CyclodextrinDocument10 pagesStability and Enzymatic Studies With Omeprazole:Hydroxypropyl - CyclodextrinPharma StpPas encore d'évaluation
- Chemical Properties and Antioxidative Activity of GlycatedDocument9 pagesChemical Properties and Antioxidative Activity of GlycatedluisbrumovskyPas encore d'évaluation
- Chicken LiverDocument10 pagesChicken LiverZoran MiladinovićPas encore d'évaluation
- Chemical Modification of A Cellulase From Aspergillus Niger: Components SystemsDocument8 pagesChemical Modification of A Cellulase From Aspergillus Niger: Components SystemsAprilia Isma DenilaPas encore d'évaluation
- Aviculare L.: Antioxidant Activity of Extract From PolygonumDocument8 pagesAviculare L.: Antioxidant Activity of Extract From PolygonumWilly YanuwarPas encore d'évaluation
- Modelling Ethanol Production From Cellulose: Separate Hydrolysis and Fermentation Versus Simultaneous Saccharification and FermentationDocument10 pagesModelling Ethanol Production From Cellulose: Separate Hydrolysis and Fermentation Versus Simultaneous Saccharification and FermentationlandolandazPas encore d'évaluation
- Cardiotoxin of The Indian Cobra (Naja Naja) Is A PyrophosphataseDocument7 pagesCardiotoxin of The Indian Cobra (Naja Naja) Is A PyrophosphataseRahul DattaPas encore d'évaluation
- Purification and Characterization of A Monoacylglycerol Lipase From The Moderately Thermophilic Bacillus Sp. H-257Document7 pagesPurification and Characterization of A Monoacylglycerol Lipase From The Moderately Thermophilic Bacillus Sp. H-257ayauPas encore d'évaluation
- 1414 431X BJMBR 1414 431X20176346Document8 pages1414 431X BJMBR 1414 431X20176346Guilherme Vasconcelos VasconcelosPas encore d'évaluation
- Shibata 2002Document6 pagesShibata 2002Hazel Kaye OcaPas encore d'évaluation
- 1977 CazzuloDocument3 pages1977 CazzuloKeyla MaydeePas encore d'évaluation
- Research Open Access: Functional Foods in Health and Disease 2011 7:232-244Document13 pagesResearch Open Access: Functional Foods in Health and Disease 2011 7:232-244Anonymous 5NXc6NuPas encore d'évaluation
- Cabana 2007Document9 pagesCabana 2007Valeria MirandaPas encore d'évaluation
- Bio-Synthesis and Hydrolysis of Ethyl Phenylacetate and Ethyl 2-Phenylpropionate in Organic Solvent by Lyophilized MyceliaDocument6 pagesBio-Synthesis and Hydrolysis of Ethyl Phenylacetate and Ethyl 2-Phenylpropionate in Organic Solvent by Lyophilized Myceliasuarezf6844Pas encore d'évaluation
- EpscDocument6 pagesEpscVohinh NgoPas encore d'évaluation
- Acetaminophen Prodrugs: Synthesis, Physicochemical Properties, and Analgesic ActivityDocument7 pagesAcetaminophen Prodrugs: Synthesis, Physicochemical Properties, and Analgesic ActivityNguyễn HàPas encore d'évaluation
- Interaction of Carbohydrates With Alcohol Dehydrogenase: Effect On Enzyme ActivityDocument5 pagesInteraction of Carbohydrates With Alcohol Dehydrogenase: Effect On Enzyme ActivityindahPas encore d'évaluation
- Forma BarataDocument2 pagesForma BarataYosel DavidPas encore d'évaluation
- Artigo ImobilizaçãoDocument6 pagesArtigo ImobilizaçãocterrasanPas encore d'évaluation
- IJFSTDocument7 pagesIJFSTElavarasan KrishnamoorthyPas encore d'évaluation
- 4 +Proline-Catalyzed+AsymmetriDocument8 pages4 +Proline-Catalyzed+Asymmetricmc107Pas encore d'évaluation
- Lee 2000Document14 pagesLee 2000Seema YadavPas encore d'évaluation
- Antioxidant Activities of Major Components of From Rice Bran Using A Linoleic Acid ModelDocument5 pagesAntioxidant Activities of Major Components of From Rice Bran Using A Linoleic Acid ModelNur SabitriPas encore d'évaluation
- Factors Affecting Captopril Stability in Aqueous SolutionDocument8 pagesFactors Affecting Captopril Stability in Aqueous SolutionTuấn HoàngPas encore d'évaluation
- AminofostineDocument5 pagesAminofostinePedro JuarezPas encore d'évaluation
- Jae PaperDocument7 pagesJae PaperanyasijoPas encore d'évaluation
- 4 RNP 0907 125Document11 pages4 RNP 0907 125Soumen ChoudhuryPas encore d'évaluation
- Chapter 4Document19 pagesChapter 4ElenaDinculescuPas encore d'évaluation
- Energy Conservation by Succinate Decarboxylation in Veillonezla PandaDocument5 pagesEnergy Conservation by Succinate Decarboxylation in Veillonezla PandaMuhamad Alim MakhrusPas encore d'évaluation
- Dendrimetos de PTDocument16 pagesDendrimetos de PTJose Miguel PalomoPas encore d'évaluation
- Paper 1Document6 pagesPaper 1Gopika SureshPas encore d'évaluation
- 10 - Chapter 6 PDFDocument19 pages10 - Chapter 6 PDFRanjith KumarPas encore d'évaluation
- Shela G., Olga M.B., Elena K., Antonin L., Nuria G.M, Ratiporn H.Document6 pagesShela G., Olga M.B., Elena K., Antonin L., Nuria G.M, Ratiporn H.Kiệt LêPas encore d'évaluation
- Cakmak1991 - Effect of Aluminium On Lipid Peroxidation, Siiperoxide DismutaseDocument7 pagesCakmak1991 - Effect of Aluminium On Lipid Peroxidation, Siiperoxide Dismutasejulie libronPas encore d'évaluation
- Park2004 PDFDocument7 pagesPark2004 PDFElson TavaresPas encore d'évaluation
- Tingkatan AntioksidanDocument5 pagesTingkatan AntioksidanMygamePas encore d'évaluation
- Huang 2013Document10 pagesHuang 2013Stefania DavidPas encore d'évaluation
- Inhibitory Protein: Effect of PhosphatasesDocument8 pagesInhibitory Protein: Effect of PhosphatasesAhdessa PerezPas encore d'évaluation
- Activity of Guaiacol Peroxidase of Solanum Lycopersicum in Presence of Detergents and Chaotropic AgentsDocument5 pagesActivity of Guaiacol Peroxidase of Solanum Lycopersicum in Presence of Detergents and Chaotropic AgentsIOSR Journal of PharmacyPas encore d'évaluation
- Gomez Et Al. 2007Document9 pagesGomez Et Al. 2007mvillarcamposPas encore d'évaluation
- 1998v46no2p401 406Document6 pages1998v46no2p401 406Kabo BakoPas encore d'évaluation
- Lago 2014Document9 pagesLago 2014Jonatas CarvalhoPas encore d'évaluation
- DPPH Radical-Scavenging Compounds From Dou-Chi, A Soybean Fermented FoodDocument0 pageDPPH Radical-Scavenging Compounds From Dou-Chi, A Soybean Fermented FoodEllen SantosPas encore d'évaluation
- Enzyme Catalytic Promiscuity The Papain-Catalyzed Knoevenagel Reaction (Paper 2)Document6 pagesEnzyme Catalytic Promiscuity The Papain-Catalyzed Knoevenagel Reaction (Paper 2)thamtusieuquayPas encore d'évaluation
- Biochemj00571 0063Document13 pagesBiochemj00571 0063KatjaVindingPas encore d'évaluation
- J. Biol. Chem.-1968-Barel-1344-8Document5 pagesJ. Biol. Chem.-1968-Barel-1344-8Nguyễn Ngô SangPas encore d'évaluation
- Erwinia Carotovora: Research ArticleDocument10 pagesErwinia Carotovora: Research Articlesrinu_aerrowPas encore d'évaluation
- Storer RJ Ferrante A 1998 Meth Mol Biol 79 81-90 chp:10.1385/0-89603-448-8:81Document10 pagesStorer RJ Ferrante A 1998 Meth Mol Biol 79 81-90 chp:10.1385/0-89603-448-8:81rjstorer9Pas encore d'évaluation
- Manuscript PBDocument33 pagesManuscript PBThirupathi Kumara Raja SelvarajPas encore d'évaluation
- GlutathioneredDocument7 pagesGlutathioneredNgot Ngao CandyPas encore d'évaluation
- Sulfonamides Determination in Chicken Meat Products From MalaysiaDocument7 pagesSulfonamides Determination in Chicken Meat Products From MalaysiaراجيرحمةربهPas encore d'évaluation
- Beta Sitosterol y GSHDocument7 pagesBeta Sitosterol y GSHEridani OlmosPas encore d'évaluation
- Zeng 1991Document10 pagesZeng 1991mfifen aristidePas encore d'évaluation
- The Role of ATP Citrate Lyase, Malic Enzyme and Fatty Acid Synthase in The Regulation of Lipid Accumulation in Cunninghamella Sp. 2A1Document6 pagesThe Role of ATP Citrate Lyase, Malic Enzyme and Fatty Acid Synthase in The Regulation of Lipid Accumulation in Cunninghamella Sp. 2A1Bara' HammadehPas encore d'évaluation
- Physician To Population Ratios Physician Supply StandardsDocument5 pagesPhysician To Population Ratios Physician Supply StandardsjackeldescargadorPas encore d'évaluation
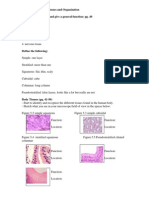
- Human Biology Laboratory 4: Body Tissues and Organization List The 4 Tissue Types and Give A General Function: Pg. 40Document4 pagesHuman Biology Laboratory 4: Body Tissues and Organization List The 4 Tissue Types and Give A General Function: Pg. 40bugyourselfPas encore d'évaluation
- Imperforate Anus PDFDocument2 pagesImperforate Anus PDFGlyneth FranciaPas encore d'évaluation
- Client Choice Plus Key FactsDocument4 pagesClient Choice Plus Key Factsjudge101Pas encore d'évaluation
- Chapter 41 - Thoracic Outlet Syndrome SynonymsDocument8 pagesChapter 41 - Thoracic Outlet Syndrome SynonymsPiero Massafra100% (1)
- Pyrolle MsdsDocument5 pagesPyrolle MsdsAmalina HazwaniPas encore d'évaluation
- Current Issues in Postoperative Pain Management PDFDocument12 pagesCurrent Issues in Postoperative Pain Management PDFikm fkunissulaPas encore d'évaluation
- Common Sports InjuryDocument71 pagesCommon Sports InjuryGlen Dizon100% (1)
- Transmission Method: What Is Pulse Oximeter?Document3 pagesTransmission Method: What Is Pulse Oximeter?samir mishraPas encore d'évaluation
- Methicillin-Resistant Staphylococcus Aureus PDFDocument23 pagesMethicillin-Resistant Staphylococcus Aureus PDFGerardo N. Pabón Gallino100% (1)
- Doctor Mi Kay Khau Nandini DhargalkarDocument51 pagesDoctor Mi Kay Khau Nandini Dhargalkarabmopalhvac0% (1)
- NICU OrganizationDocument5 pagesNICU Organizationgopscharan100% (1)
- Point of CareDocument5 pagesPoint of CareDaniela RochaPas encore d'évaluation
- Fiji Times Jan 7Document48 pagesFiji Times Jan 7fijitimescanadaPas encore d'évaluation
- Cardiac GlycosidesDocument8 pagesCardiac GlycosidesShan Sicat100% (1)
- SEE WITHOUT Glasses PreviewDocument21 pagesSEE WITHOUT Glasses PreviewHokusyPas encore d'évaluation
- Organon of Medicine All Years 2marks Question Viva Questions AnswersDocument12 pagesOrganon of Medicine All Years 2marks Question Viva Questions Answersabcxyz15021999Pas encore d'évaluation
- 7 Uses For Medical MarijuanaDocument4 pages7 Uses For Medical MarijuanaYhojan Surco ZavalaPas encore d'évaluation
- Concept Map 3 MM (Abdominal Pain)Document2 pagesConcept Map 3 MM (Abdominal Pain)Matt McKinleyPas encore d'évaluation
- Periop QuizesDocument9 pagesPeriop QuizesAnonymous ZQ4gHahzPas encore d'évaluation
- Raw Materials & Purification - NitrogenDocument9 pagesRaw Materials & Purification - Nitrogensexyrusty0% (1)
- Herbal Supplements-Drug Interactions Lam Huang Hall (CRC 2006) BBS PDFDocument330 pagesHerbal Supplements-Drug Interactions Lam Huang Hall (CRC 2006) BBS PDFAnonymous m4A4Yuvrwm100% (1)
- A Case Study On Ascites of Hepatic OriginDocument4 pagesA Case Study On Ascites of Hepatic OriginFaisal MohommadPas encore d'évaluation
- Psychology From Islamic Perspective (Amber)Document22 pagesPsychology From Islamic Perspective (Amber)Rðbið HðquePas encore d'évaluation
- Microscopic Examination of UrineDocument89 pagesMicroscopic Examination of UrineLucia SurduPas encore d'évaluation
- Choking and AspirationDocument23 pagesChoking and AspirationAgni MubarokPas encore d'évaluation
- Limfoma Kutis Pada Pasien Yang Semula Di DiagnosisDocument34 pagesLimfoma Kutis Pada Pasien Yang Semula Di DiagnosisChris MulyoPas encore d'évaluation
- Legal MedDocument42 pagesLegal Medaisajane_rmt100% (6)
- Acute Limb Ischemia: Clinical PracticeDocument9 pagesAcute Limb Ischemia: Clinical PracticeIndah MaulidawatiPas encore d'évaluation
- Massive Transfusion ProtocolDocument2 pagesMassive Transfusion ProtocolmukriPas encore d'évaluation