Vous aimerez peut-être aussi
- Quinolone Derivatives As Antitubercular Drugs: Medicinal Chemistry ResearchDocument14 pagesQuinolone Derivatives As Antitubercular Drugs: Medicinal Chemistry ResearchArasy PratamaPas encore d'évaluation
- Safari - 17 Okt 2019 14.41Document1 pageSafari - 17 Okt 2019 14.41D MayaPas encore d'évaluation
- Berry-2004-Journal of NeurochemistryDocument15 pagesBerry-2004-Journal of NeurochemistryGuadalupe BautistaPas encore d'évaluation
- 3 ANSDrugsDocument75 pages3 ANSDrugsLinearPas encore d'évaluation
- roledopamineOMCL2016 9730467Document13 pagesroledopamineOMCL2016 9730467gbautista.fuertePas encore d'évaluation
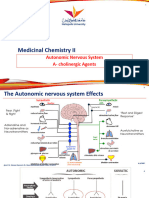
- Drugs Acting On ANSDocument28 pagesDrugs Acting On ANSVidhi MehtaPas encore d'évaluation
- Phosphate Group (Double-Ringed: DeoxyriboseDocument6 pagesPhosphate Group (Double-Ringed: Deoxyriboseabdulkareemfarida4Pas encore d'évaluation
- Central Nervous System Finals 2ndyrDocument7 pagesCentral Nervous System Finals 2ndyrkassy yeonPas encore d'évaluation
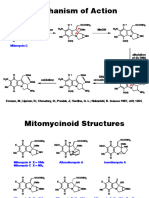
- BatzelladineDocument13 pagesBatzelladinebroccolikimPas encore d'évaluation
- 11TMDocument5 pages11TMTho PhanPas encore d'évaluation
- Cartel ACSDocument1 pageCartel ACSHua LinPas encore d'évaluation
- 573 PDFDocument7 pages573 PDFAnkit Kumar VermaPas encore d'évaluation
- Histamine 1Document46 pagesHistamine 1raviPas encore d'évaluation
- Davis MDCH 5210 - Anticonvulsants 2006Document19 pagesDavis MDCH 5210 - Anticonvulsants 2006Michael ThomPas encore d'évaluation
- Fundamentals of Med Chem 2024 - Student SldiesDocument46 pagesFundamentals of Med Chem 2024 - Student SldiesMartina DiazPas encore d'évaluation
- Tridentate Hydrazone Metal Complexes Derived From Cephalexinand 2-Hydrazinopyridine Synthesis, Characterization and Antibacterial ActivityDocument7 pagesTridentate Hydrazone Metal Complexes Derived From Cephalexinand 2-Hydrazinopyridine Synthesis, Characterization and Antibacterial ActivityGina DobritescuPas encore d'évaluation
- Nmda PDFDocument25 pagesNmda PDFMutescuAnamariaPas encore d'évaluation
- The β - D Glucose Scaffold as a β- Turn Mimetic: Saidulu DaraDocument19 pagesThe β - D Glucose Scaffold as a β- Turn Mimetic: Saidulu Daraglreddy09Pas encore d'évaluation
- FULL PAPER1 Synthesis of Pyridazin-3 (2H) - One Derivatives by The Reaction of CH-acids With DichlorodiazadienesDocument10 pagesFULL PAPER1 Synthesis of Pyridazin-3 (2H) - One Derivatives by The Reaction of CH-acids With DichlorodiazadienesHamza AssilaPas encore d'évaluation
- Central Nervous System StimulantsDocument10 pagesCentral Nervous System StimulantsCristian F ApontePas encore d'évaluation
- Product Information: MethylcobalaminDocument1 pageProduct Information: MethylcobalaminVandit KharePas encore d'évaluation
- Amine NeurotransmittersDocument7 pagesAmine NeurotransmittersFrancisco BecerraPas encore d'évaluation
- A Fatal Intoxication Following The Ingestion of 5-Methoxy-N, N-Dimethyltryptamine in An Ayahuasca PreparationDocument4 pagesA Fatal Intoxication Following The Ingestion of 5-Methoxy-N, N-Dimethyltryptamine in An Ayahuasca PreparationpavelPas encore d'évaluation
- A Facile and e Cient Synthesis of Highly Functionalised 3,3 - Dispiropyrrolidine-And 3,3 - Dispiropyrrolizidine Bisoxindoles Via (3+2) CycloadditionDocument5 pagesA Facile and e Cient Synthesis of Highly Functionalised 3,3 - Dispiropyrrolidine-And 3,3 - Dispiropyrrolizidine Bisoxindoles Via (3+2) CycloadditionHamed HamidpourPas encore d'évaluation
- Adrenergic Agents SeminarDocument48 pagesAdrenergic Agents SeminarTheBoss 20Pas encore d'évaluation
- Alcalolides de Triptófano Final Parte 1Document43 pagesAlcalolides de Triptófano Final Parte 1JUAN JOSE OLMOS CAMPOPas encore d'évaluation
- Alcalolides de Triptófano Final Parte 1Document43 pagesAlcalolides de Triptófano Final Parte 1MANUELA RAMIREZ SEPULVEDAPas encore d'évaluation
- Non Mendelian GeneticsDocument48 pagesNon Mendelian GeneticsFrank Louie MagadiaPas encore d'évaluation
- Samar DCU Poster 2007Document1 pageSamar DCU Poster 2007Zainab altafPas encore d'évaluation
- Cao2013 Synthesis and Structureeactivity Relationships of Harmine Derivatives As PotentialDocument9 pagesCao2013 Synthesis and Structureeactivity Relationships of Harmine Derivatives As Potentialtaoufik akabliPas encore d'évaluation
- RJS - Volume 28 - Issue 2 - Pages 146-151Document6 pagesRJS - Volume 28 - Issue 2 - Pages 146-151Shubham BendkulePas encore d'évaluation
- 1 Lec 1-Parasympathetic AgentsDocument33 pages1 Lec 1-Parasympathetic Agentsabdelrahmanmosleh75Pas encore d'évaluation
- Powerpoint The Language of LifeDocument59 pagesPowerpoint The Language of LifescienceandapologeticsPas encore d'évaluation
- Wien 06 05 From Leads To DrugsDocument20 pagesWien 06 05 From Leads To DrugsSaeful AminPas encore d'évaluation
- 7 1 Andrzeje-331 17-26Document9 pages7 1 Andrzeje-331 17-26Mia L. AyuningtyasPas encore d'évaluation
- Synthesis, Characterization, and Antitumor Activity of New Copper (I) and Mercury (II) ComplexesDocument8 pagesSynthesis, Characterization, and Antitumor Activity of New Copper (I) and Mercury (II) ComplexesRoman RusnacPas encore d'évaluation
- Chapter 9 PDFDocument41 pagesChapter 9 PDFIziz De La CruzPas encore d'évaluation
- EJCHEM - Volume 64 - Issue 7 - Pages 3791-3800Document10 pagesEJCHEM - Volume 64 - Issue 7 - Pages 3791-3800Popusoi AnaPas encore d'évaluation
- A Fatal Intoxication Following The Ingestion of 5-Methoxy-N, N-Dimethyltryptamine in An Ayahuasca PreparationDocument4 pagesA Fatal Intoxication Following The Ingestion of 5-Methoxy-N, N-Dimethyltryptamine in An Ayahuasca PreparationemilioPas encore d'évaluation
- 4 Med ChemDocument2 pages4 Med ChemsupriyoPas encore d'évaluation
- D Etermination of N, N-Dimethyltryptamine and B-Carboline Alkaloids in Human Plasma Following Oral Administration of AyahuascaDocument11 pagesD Etermination of N, N-Dimethyltryptamine and B-Carboline Alkaloids in Human Plasma Following Oral Administration of AyahuascaoolonPas encore d'évaluation
- Concise SynthesisDocument6 pagesConcise SynthesisAnh Pham Le NgocPas encore d'évaluation
- Jasplakinolide, Jaspis Johnstoni: Tel 0800 6931 000 Tel (800) 628-8470 Tel 0115 9430 840Document2 pagesJasplakinolide, Jaspis Johnstoni: Tel 0800 6931 000 Tel (800) 628-8470 Tel 0115 9430 840Raymond BaldelovarPas encore d'évaluation
- A One-Pot Transition-Metal-Free Tandem Process To 1,4-Benzodiazepine ScaffoldsDocument7 pagesA One-Pot Transition-Metal-Free Tandem Process To 1,4-Benzodiazepine ScaffoldsNoimurPas encore d'évaluation
- HallucinogensDocument9 pagesHallucinogensMada madalinaPas encore d'évaluation
- Lec14 PDFDocument11 pagesLec14 PDFAishwarya MankaniPas encore d'évaluation
- BA MS Fingerprinting PDFDocument7 pagesBA MS Fingerprinting PDFIon VasilescuPas encore d'évaluation
- Creation of A Long-Acting Nanoformulated DolutegraDocument14 pagesCreation of A Long-Acting Nanoformulated DolutegraRanga NdhlovuPas encore d'évaluation
- C 7 NJ 04648 KDocument15 pagesC 7 NJ 04648 Kmamut343Pas encore d'évaluation
- Antineoplastic: Kimia Medisinal S-1 Farmasi Iik BWDocument61 pagesAntineoplastic: Kimia Medisinal S-1 Farmasi Iik BWKiki KartikaPas encore d'évaluation
- s44160-022-00080-x SEMINARIODocument6 pagess44160-022-00080-x SEMINARIOMarcos GPas encore d'évaluation
- Tugas Michelle 2Document7 pagesTugas Michelle 2andre nicholasPas encore d'évaluation
- Total Sintesis1Document34 pagesTotal Sintesis1M Irhash ShalihinPas encore d'évaluation
- Research ArticleDocument8 pagesResearch ArticleNandhy AgusPas encore d'évaluation
- Drug Design: Drug Structure & Pharmacological ActivityDocument55 pagesDrug Design: Drug Structure & Pharmacological ActivityMominul IslamPas encore d'évaluation
- Cholinergics Part-IIIDocument35 pagesCholinergics Part-IIIYaman Adnan QabajaPas encore d'évaluation
- Scipharm 86 00031Document21 pagesScipharm 86 00031aruna aPas encore d'évaluation
- 2018 HarmineDocument15 pages2018 Harminetaoufik akabliPas encore d'évaluation
- Treatment of Benzodiazepine DependenceDocument11 pagesTreatment of Benzodiazepine DependenceRoberto López Mata0% (1)
- 1397044565-Broto Santoso-Ums Perhipba 2014 3Document14 pages1397044565-Broto Santoso-Ums Perhipba 2014 3Dita MarthariniPas encore d'évaluation
- Jain2012 PDFDocument13 pagesJain2012 PDFDita MarthariniPas encore d'évaluation
- Photochemical Synthesis of Gold Nanoparticles by The Sunlight Radiation Using A Seeding ApproachDocument9 pagesPhotochemical Synthesis of Gold Nanoparticles by The Sunlight Radiation Using A Seeding ApproachDita MarthariniPas encore d'évaluation
- Kandidat Skripsi Fakultas Kedokteran Universitas Sam Ratulangi Manado Bagian Kardiologi Fakultas Kedokteran Universitas Sam Ratulangi ManadoDocument11 pagesKandidat Skripsi Fakultas Kedokteran Universitas Sam Ratulangi Manado Bagian Kardiologi Fakultas Kedokteran Universitas Sam Ratulangi ManadoDita MarthariniPas encore d'évaluation
- 1,2 JugaDocument7 pages1,2 JugaDita MarthariniPas encore d'évaluation
- Tuesday 19 May 2020: Biology A (Salters Nuffield)Document24 pagesTuesday 19 May 2020: Biology A (Salters Nuffield)Mariam HasanPas encore d'évaluation
- DR Abdul Aziz Mohamed YusoffDocument6 pagesDR Abdul Aziz Mohamed YusoffAhriShaarmineeIIPas encore d'évaluation
- Introduction To PathologyDocument21 pagesIntroduction To PathologylavanyagunduPas encore d'évaluation
- ChemistrySelect 2017Document7 pagesChemistrySelect 2017Sandeep GargPas encore d'évaluation
- Biochemistry: A Short CourseDocument31 pagesBiochemistry: A Short CourseEli JohnsonPas encore d'évaluation
- Milk ProteinsDocument5 pagesMilk ProteinsFandy RoevPas encore d'évaluation
- Physiology - LipidsDocument45 pagesPhysiology - LipidsMark Albert Concepcion NardoPas encore d'évaluation
- CHEM41290 Notes Part IDocument25 pagesCHEM41290 Notes Part IJulia MachajPas encore d'évaluation
- Cloning Vector: BY: Dr. Mukesh Kumar BhardwajDocument23 pagesCloning Vector: BY: Dr. Mukesh Kumar Bhardwajمحمود محمدPas encore d'évaluation
- Protein Requirements and Supplementation in Strength Sports: Nutrition July 2004Document8 pagesProtein Requirements and Supplementation in Strength Sports: Nutrition July 2004Andrei ChirilaPas encore d'évaluation
- BCH 261 Midterm 2015 MTCDocument3 pagesBCH 261 Midterm 2015 MTCbecca0% (1)
- Genetic EngineeringDocument39 pagesGenetic EngineeringAerone C AntolinPas encore d'évaluation
- Chemistry of Life: Name: Jose David Parada Polentino Date: 08/10/2020Document1 pageChemistry of Life: Name: Jose David Parada Polentino Date: 08/10/2020David PolentinoPas encore d'évaluation
- Molecular ConnectionDocument4 pagesMolecular ConnectionCaryl Louise ParlanPas encore d'évaluation
- Chemistry Chemical EngineeringDocument124 pagesChemistry Chemical Engineeringjrobs314Pas encore d'évaluation
- Diet Effects On MitochondriaDocument26 pagesDiet Effects On MitochondriabenZen100% (1)
- Structural Components of The Cell MembraneDocument2 pagesStructural Components of The Cell MembraneYumiPas encore d'évaluation
- Intestinal and Colonic Physiology and Biochemistry (Part 2 - Secretion, Digestion, Absorption) (Slides With Notes)Document69 pagesIntestinal and Colonic Physiology and Biochemistry (Part 2 - Secretion, Digestion, Absorption) (Slides With Notes)kuchipuchi12Pas encore d'évaluation
- Proteins (Lec)Document3 pagesProteins (Lec)Dashi MeiPas encore d'évaluation
- Paternity Test LabDocument4 pagesPaternity Test LabChase HildebrandPas encore d'évaluation
- Genetically Modified OrganismsDocument7 pagesGenetically Modified OrganismsPrajwal PatilPas encore d'évaluation
- 100 MCAT TipsDocument23 pages100 MCAT TipsJhilianne Batino100% (1)
- Cell MembraneDocument5 pagesCell MembraneJenica CruzPas encore d'évaluation
- SLG Chem 3 LG 5.3 Structure of Amino Acids and Levels of Protein StructureDocument9 pagesSLG Chem 3 LG 5.3 Structure of Amino Acids and Levels of Protein StructurefranzachilleslindayagPas encore d'évaluation
- Chap 003 Anatomy and PhysiologyDocument43 pagesChap 003 Anatomy and PhysiologyahmshiPas encore d'évaluation
- SOP 3.04 DNA Extraction From BloodDocument4 pagesSOP 3.04 DNA Extraction From BloodMeetali GuptaPas encore d'évaluation
- Carbohydrate Part 1Document29 pagesCarbohydrate Part 1Des LumabanPas encore d'évaluation
- Test Bank For The Biology of Cancer Second EditionDocument7 pagesTest Bank For The Biology of Cancer Second EditionDale Passe100% (26)
- Bharti 1927Document19 pagesBharti 1927vikasPas encore d'évaluation
- Monomer - WikipediaDocument3 pagesMonomer - Wikipediaramthecharm_46098467Pas encore d'évaluation