Vous aimerez peut-être aussi
- (Fortschritte Der Chemie Organischer Naturstoffe - Progress in The Chemistry of Organic Natural Products 81) S. Huneck (Auth.), Prof. W. Herz, Prof. H. Falk, Prof. G. W. Kirby, Prof. R. E. Moore (Eds.Document320 pages(Fortschritte Der Chemie Organischer Naturstoffe - Progress in The Chemistry of Organic Natural Products 81) S. Huneck (Auth.), Prof. W. Herz, Prof. H. Falk, Prof. G. W. Kirby, Prof. R. E. Moore (Eds.vita100% (1)
- Gdlhub GDL Res 2014 Studiawanh 36034 11. Daft ADocument2 pagesGdlhub GDL Res 2014 Studiawanh 36034 11. Daft AvitaPas encore d'évaluation
- Bir Bahadur, Manchikatla Venkat Rajam, Leela Sahijram, K.v. Krishnamurthy (Eds.)-Plant Biology and Biotechnology_ Volume I_ Plant Diversity, Organization, Function and Improvement-Springer India (2015Document831 pagesBir Bahadur, Manchikatla Venkat Rajam, Leela Sahijram, K.v. Krishnamurthy (Eds.)-Plant Biology and Biotechnology_ Volume I_ Plant Diversity, Organization, Function and Improvement-Springer India (2015vita100% (1)
- Paper 08Document8 pagesPaper 08vitaPas encore d'évaluation
- Baseline Element Composition of Foliose and Fruticose Lichens Along The Steep Climatic Gradient of SW Patagonia (Aisen Region, Chile)Document9 pagesBaseline Element Composition of Foliose and Fruticose Lichens Along The Steep Climatic Gradient of SW Patagonia (Aisen Region, Chile)vitaPas encore d'évaluation
- Penicillium Citrinum FNCC 6111) and Agar Diffusion Method. Five Isolates WithDocument1 pagePenicillium Citrinum FNCC 6111) and Agar Diffusion Method. Five Isolates WithvitaPas encore d'évaluation
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5795)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Lesson Plan: Instructor: Date Topic: Grade Level: Subject ObjectivesDocument5 pagesLesson Plan: Instructor: Date Topic: Grade Level: Subject Objectivesapi-340265930Pas encore d'évaluation
- The Structure of Deception: Validation of The Lying Profile QuestionnaireDocument16 pagesThe Structure of Deception: Validation of The Lying Profile QuestionnaireNancy DrewPas encore d'évaluation
- Capillary PressureDocument12 pagesCapillary PressureamahaminerPas encore d'évaluation
- Mathematics ExcerciseDocument74 pagesMathematics Excerciseserge129100% (1)
- Multithreading AlgorithmsDocument36 pagesMultithreading AlgorithmsAsna TariqPas encore d'évaluation
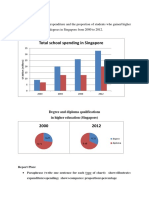
- Total School Spending in Singapore: Task 1 020219Document6 pagesTotal School Spending in Singapore: Task 1 020219viper_4554Pas encore d'évaluation
- SLC 10 ScienceDocument16 pagesSLC 10 SciencePffflyers KurnawanPas encore d'évaluation
- Lesson 2 (Locating Main Ideas)Document18 pagesLesson 2 (Locating Main Ideas)Glyneth Dela TorrePas encore d'évaluation
- Tcps 2 Final WebDocument218 pagesTcps 2 Final WebBornaGhannadiPas encore d'évaluation
- HS ĐỀ CƯƠNG KIỂM TRA GIỮA KỲ I KHOI 10 NH 23 24Document6 pagesHS ĐỀ CƯƠNG KIỂM TRA GIỮA KỲ I KHOI 10 NH 23 24Ngô Hiểu NhiênPas encore d'évaluation
- IBRO News 2004Document8 pagesIBRO News 2004International Brain Research Organization100% (1)
- Heat and Cooling LoadDocument2 pagesHeat and Cooling LoadAshokPas encore d'évaluation
- Gender InequalityDocument9 pagesGender InequalityRhzPas encore d'évaluation
- 02.03.2017 SR Mechanical Engineer Externe Vacature InnoluxDocument1 page02.03.2017 SR Mechanical Engineer Externe Vacature InnoluxMichel van WordragenPas encore d'évaluation
- Proposal Defense Form Chapter 1 3Document3 pagesProposal Defense Form Chapter 1 3Jason BinondoPas encore d'évaluation
- OS Lab ManualDocument47 pagesOS Lab ManualNivedhithaVPas encore d'évaluation
- Behavan00004 0003Document19 pagesBehavan00004 0003Swati GroverPas encore d'évaluation
- Chapter 17Document78 pagesChapter 17Romar PanopioPas encore d'évaluation
- Журнал JournalistDocument36 pagesЖурнал JournalistSofia SorochinskaiaPas encore d'évaluation
- Design BasisDocument9 pagesDesign BasisSamiksha RajdevPas encore d'évaluation
- Waves Scattering Induced by An Interface Crack in A Coated MaterialDocument6 pagesWaves Scattering Induced by An Interface Crack in A Coated MaterialDarioLampaPas encore d'évaluation
- My Home, My School: Meet and GreetDocument2 pagesMy Home, My School: Meet and GreetAcebuque Arby Immanuel GopeteoPas encore d'évaluation
- Lesson Plan ComsumerismDocument6 pagesLesson Plan ComsumerismMan Eugenia50% (4)
- Critical Point of View: A Wikipedia ReaderDocument195 pagesCritical Point of View: A Wikipedia Readerrll307Pas encore d'évaluation
- Cubic FunctionsDocument9 pagesCubic FunctionsShrey JainPas encore d'évaluation
- DSpace Configuration PDFDocument1 pageDSpace Configuration PDFArshad SanwalPas encore d'évaluation
- Document 1 PetDocument9 pagesDocument 1 PetRoel Jr Pinaroc DolaypanPas encore d'évaluation
- Project PlanDocument8 pagesProject PlanRodsheen100% (1)
- Sika Bro - E - Sikadur-Combiflex SG System - High Performance Joint - Crack Waterproofing System (09.2010)Document8 pagesSika Bro - E - Sikadur-Combiflex SG System - High Performance Joint - Crack Waterproofing System (09.2010)Joh SongthamPas encore d'évaluation
- 1 SMDocument9 pages1 SMSelly ManaluPas encore d'évaluation