Vous aimerez peut-être aussi
- HulaDocument19 pagesHulaJon LenonPas encore d'évaluation
- HulaDocument19 pagesHulaJon LenonPas encore d'évaluation
- HulaDocument19 pagesHulaJon LenonPas encore d'évaluation
- HulaDocument19 pagesHulaJon LenonPas encore d'évaluation
- HulaDocument19 pagesHulaJon LenonPas encore d'évaluation
- HulaDocument19 pagesHulaJon LenonPas encore d'évaluation
- HulaDocument21 pagesHulaJon LenonPas encore d'évaluation
- HulaDocument19 pagesHulaJon LenonPas encore d'évaluation
- HulaDocument8 pagesHulaJon LenonPas encore d'évaluation
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (120)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- PS Ebook BagsPouches2 1116 HRDocument58 pagesPS Ebook BagsPouches2 1116 HRGovindarajulu RPas encore d'évaluation
- Ujian Diagnostik Ting 2 EnglishDocument9 pagesUjian Diagnostik Ting 2 EnglishJ-Gie JaulahPas encore d'évaluation
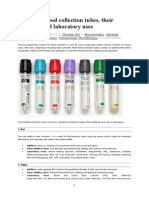
- Eprubete. Culori. UtilizareDocument3 pagesEprubete. Culori. UtilizareCuCUPas encore d'évaluation
- Chapter 2Document5 pagesChapter 2ERICKA MAE NATOPas encore d'évaluation
- Components of A Rainwater Harvesting SystemDocument38 pagesComponents of A Rainwater Harvesting SystembudiperPas encore d'évaluation
- User's Manual Eco Friendly Park BenchDocument35 pagesUser's Manual Eco Friendly Park BenchAmirulHanif AlyahyaPas encore d'évaluation
- Wetted Wall Gas AbsorptionDocument9 pagesWetted Wall Gas AbsorptionSiraj AL sharifPas encore d'évaluation
- APPLE FRITTER CAKE Cook Heavenly RecipesDocument4 pagesAPPLE FRITTER CAKE Cook Heavenly Recipes1940LaSallePas encore d'évaluation
- X FEDEX EIDocument13 pagesX FEDEX EINISREEN WAYAPas encore d'évaluation
- Laboratory Diagnosis of Parasitic DiseasesDocument57 pagesLaboratory Diagnosis of Parasitic DiseasesAmanuel MaruPas encore d'évaluation
- Electrical Rooms Fire FightingDocument2 pagesElectrical Rooms Fire Fightingashraf saidPas encore d'évaluation
- WAUZZZ8K0BA159120Document10 pagesWAUZZZ8K0BA159120Vedad VedaddPas encore d'évaluation
- Types of Business OwnershipDocument2 pagesTypes of Business Ownershipprelovedseller bnPas encore d'évaluation
- Rodents and Sectional Title ComplexDocument2 pagesRodents and Sectional Title ComplexFredSmith777Pas encore d'évaluation
- Pia AlgebraDocument12 pagesPia AlgebraCarvajal EdithPas encore d'évaluation
- Factorial Dimensions of Employee Engagement in Public and Private Sector BanksDocument5 pagesFactorial Dimensions of Employee Engagement in Public and Private Sector BanksCma Pushparaj KulkarniPas encore d'évaluation
- Music Therapy: Treatment For Grade 11 Stem Students Who Suffer Stress From Basic CalculusDocument12 pagesMusic Therapy: Treatment For Grade 11 Stem Students Who Suffer Stress From Basic CalculusArvinel L. VileganoPas encore d'évaluation
- Bandura Social Cognitive TheoryDocument93 pagesBandura Social Cognitive TheoryВикторияPas encore d'évaluation
- How To Make Panna CottaDocument3 pagesHow To Make Panna CottaBintPas encore d'évaluation
- Separating Mixtures: Techniques and Applications: Evaporation, Distillation and FiltrationDocument4 pagesSeparating Mixtures: Techniques and Applications: Evaporation, Distillation and FiltrationAndrea SobredillaPas encore d'évaluation
- MS Fresher HR DocumentDocument4 pagesMS Fresher HR DocumentJahanvi KambojPas encore d'évaluation
- 2457-Article Text-14907-2-10-20120724Document6 pages2457-Article Text-14907-2-10-20120724desi meleniaPas encore d'évaluation
- P13Document21 pagesP13Saeful AzizPas encore d'évaluation
- Cardiac AmyloidosisDocument3 pagesCardiac AmyloidosisPrasad SrbPas encore d'évaluation
- FALLSEM2021-22 EEE3004 ETH VL2021220100810 Reference Material I 16-Aug-2021 7 Controlled Rectifier NewDocument51 pagesFALLSEM2021-22 EEE3004 ETH VL2021220100810 Reference Material I 16-Aug-2021 7 Controlled Rectifier NewVAHEESPas encore d'évaluation
- LISD LetterDocument2 pagesLISD LetterAnonymous Pb39klJ100% (1)
- SubstationsDocument39 pagesSubstationspPas encore d'évaluation
- Les Essences D'amelie BrochureDocument8 pagesLes Essences D'amelie BrochuresayonarasPas encore d'évaluation
- Hydraulic Fluid CategoriesDocument3 pagesHydraulic Fluid CategoriesJako MishyPas encore d'évaluation
- Vishaka GuidelinesDocument4 pagesVishaka GuidelinesAakashKumarPas encore d'évaluation