Vous aimerez peut-être aussi
- Neurobiology of Sleep and MemoryD'EverandNeurobiology of Sleep and MemoryJames McGaughPas encore d'évaluation
- Serotonin Neurons, Neuroplasticity, and Homeostasis of Neural TissueDocument13 pagesSerotonin Neurons, Neuroplasticity, and Homeostasis of Neural TissueFuyumi RiaPas encore d'évaluation
- Extra Credit Neuroscience TwoDocument3 pagesExtra Credit Neuroscience TwoCaroline SummerPas encore d'évaluation
- The Neural Mechanisms of Gustation: A Distributed Processing CodeDocument12 pagesThe Neural Mechanisms of Gustation: A Distributed Processing CodesylPas encore d'évaluation
- Morphological and Functional Diversity of First-Order Somatosensory NeuronsDocument10 pagesMorphological and Functional Diversity of First-Order Somatosensory NeuronsgiorgiocrucittiPas encore d'évaluation
- Sleep, Emotions, and Visceral Control: I. N. Pigarev and M. L. PigarevaDocument12 pagesSleep, Emotions, and Visceral Control: I. N. Pigarev and M. L. PigarevaMariaPas encore d'évaluation
- Walsh 2000 PDFDocument7 pagesWalsh 2000 PDFChrysoula GkaniPas encore d'évaluation
- 2008 - Ehninger, Kempermann - Neurogenesis in The Adult HippocampusDocument8 pages2008 - Ehninger, Kempermann - Neurogenesis in The Adult HippocampusNityananda PortelladaPas encore d'évaluation
- Generalized Potential of Adult Neural Stem Cells: EportsDocument5 pagesGeneralized Potential of Adult Neural Stem Cells: EportsmisterxPas encore d'évaluation
- Currie, 2016 PDFDocument18 pagesCurrie, 2016 PDF22gaefPas encore d'évaluation
- Plant Development: Brassinosteroids Go Out of Bounds: PTN 1 PTN 2 ADocument3 pagesPlant Development: Brassinosteroids Go Out of Bounds: PTN 1 PTN 2 AAle LuPas encore d'évaluation
- Papanicolaou - Brain Mechanisms For Reading in Children With and Without Dyslexia A Review of Studies of Normal Development and PlasticityDocument21 pagesPapanicolaou - Brain Mechanisms For Reading in Children With and Without Dyslexia A Review of Studies of Normal Development and PlasticityBrittany Caroline BlackPas encore d'évaluation
- Review Article TekuDocument9 pagesReview Article Tekushubhamatilkar04Pas encore d'évaluation
- Neurogenese AdultosDocument17 pagesNeurogenese AdultosDenise GarciaPas encore d'évaluation
- Sleep Memory Processing The Sequential Hypothesis (Theoretical Lens)Document8 pagesSleep Memory Processing The Sequential Hypothesis (Theoretical Lens)Sebastian CrayPas encore d'évaluation
- Study of Neurosecretory Cells in OdonataDocument16 pagesStudy of Neurosecretory Cells in Odonatashubhamatilkar04Pas encore d'évaluation
- Patzke 2013Document23 pagesPatzke 2013Kamilla souzaPas encore d'évaluation
- The Functional Utility of Consciousness Depends On Content As Well As On StateDocument73 pagesThe Functional Utility of Consciousness Depends On Content As Well As On StatePusat Bahasa NasionalPas encore d'évaluation
- Organelos en Sueño PDFDocument14 pagesOrganelos en Sueño PDFLEISLY TATIANA OVIEDO GOMEZPas encore d'évaluation
- 幹細胞與前瞻Document14 pages幹細胞與前瞻lunamedinaPas encore d'évaluation
- References: Section I / GeneralDocument10 pagesReferences: Section I / GeneralAndreea LăzăroiuPas encore d'évaluation
- Posner Attentional NetworksDocument5 pagesPosner Attentional NetworksBarnabyDPas encore d'évaluation
- Cognitive Map NobelDocument2 pagesCognitive Map NobelSakditad SaowapaPas encore d'évaluation
- Pain and Its Control in R 2018 Veterinary Clinics of North America Exotic ADocument16 pagesPain and Its Control in R 2018 Veterinary Clinics of North America Exotic AYaiza Garcia CasadoPas encore d'évaluation
- Thalamus Modulates Consciousness Via Layer Specific Control of Cortex Michelle J Redinbaugh Jessica M Phillips Niranjan A Kambi Sounak Mohanta Samantha Andryk Gaven L Full ChapterDocument46 pagesThalamus Modulates Consciousness Via Layer Specific Control of Cortex Michelle J Redinbaugh Jessica M Phillips Niranjan A Kambi Sounak Mohanta Samantha Andryk Gaven L Full Chapterronald.kenworthy366100% (7)
- Thalamic Neuron TheoryDocument18 pagesThalamic Neuron TheorybazediPas encore d'évaluation
- Types of Neurons in The Enteric Nervous SystemDocument10 pagesTypes of Neurons in The Enteric Nervous SystemErnesto Ochoa MonroyPas encore d'évaluation
- Miller2012 Braqin TeasersDocument1 pageMiller2012 Braqin Teasersiulia andreeaPas encore d'évaluation
- Neuroanatomic Connectivity of The Human Ascending ArousalDocument16 pagesNeuroanatomic Connectivity of The Human Ascending ArousalBritany SantanaPas encore d'évaluation
- Evidence For A Life-Span Theory of Socioemotional SelectivityDocument7 pagesEvidence For A Life-Span Theory of Socioemotional SelectivityNadia Oyarzo MillarPas encore d'évaluation
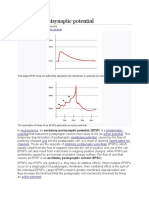
- Excitatory Postsynaptic Potential: Jump To Navigation Jump To SearchDocument6 pagesExcitatory Postsynaptic Potential: Jump To Navigation Jump To SearchJennie KimPas encore d'évaluation
- Mr. Phinoj K Abraham (Moth) : Assistant Professor, SRM College of Occupational TherapyDocument34 pagesMr. Phinoj K Abraham (Moth) : Assistant Professor, SRM College of Occupational TherapyKazi AijazPas encore d'évaluation
- Cereb. Cortex 2003 Duan 950 61Document12 pagesCereb. Cortex 2003 Duan 950 61AlfonsoVillarroelCalistoPas encore d'évaluation
- Autonomic NeuroscienceDocument13 pagesAutonomic NeuroscienceReila KnbsPas encore d'évaluation
- Sonic HedgehogDocument11 pagesSonic HedgehogPaola Rivera ÁvilaPas encore d'évaluation
- Referencia 39 2021 Kitazawa Mutacion Adrb1-A187vDocument20 pagesReferencia 39 2021 Kitazawa Mutacion Adrb1-A187vKaren RoblesPas encore d'évaluation
- MicrosDocument11 pagesMicrosEka JuliantaraPas encore d'évaluation
- Journal Pbio 3002242Document37 pagesJournal Pbio 3002242Raymundo Lopez NPas encore d'évaluation
- Gallup Contagious Yawning Systematic ReviewDocument13 pagesGallup Contagious Yawning Systematic ReviewShaelynPas encore d'évaluation
- Ajbsr MS Id 000960Document14 pagesAjbsr MS Id 000960TrishaPas encore d'évaluation
- Natural Waking and Sleep State A View From Inside Neocortical NeuronsDocument17 pagesNatural Waking and Sleep State A View From Inside Neocortical NeuronsLeslie AcevesPas encore d'évaluation
- Sleep 1Document40 pagesSleep 1Sst YadavPas encore d'évaluation
- Homer1a Is A Core Brain Molecular Correlate of Sleep LossDocument6 pagesHomer1a Is A Core Brain Molecular Correlate of Sleep LosswestPas encore d'évaluation
- Blume Et Al - 2018 - Standing Sentinel During Human SleepDocument11 pagesBlume Et Al - 2018 - Standing Sentinel During Human SleepChandan ShandilPas encore d'évaluation
- Vivar 2016 Running Rewires The Neuronal Network of Adult-Born DentateDocument14 pagesVivar 2016 Running Rewires The Neuronal Network of Adult-Born DentateJosé Luis García MezaPas encore d'évaluation
- Latest Abstract and Researches About MotivationDocument2 pagesLatest Abstract and Researches About MotivationJoymi ClairePas encore d'évaluation
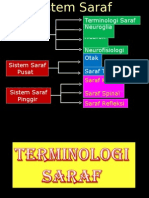
- Sistem Saraf 2Document7 pagesSistem Saraf 2Neza SepteviaPas encore d'évaluation
- The Midbrain Periaqueductal Gray As An Integrative and Interoceptive Neural Structure For BreathingDocument10 pagesThe Midbrain Periaqueductal Gray As An Integrative and Interoceptive Neural Structure For BreathingAlessandra LacerdaPas encore d'évaluation
- Synapse and MemoryDocument19 pagesSynapse and Memorysomya mathurPas encore d'évaluation
- Forebrain Engraftment by Human Glial Progenitor Cells Enhances Synaptic Plasticity and Learning in Adult MiceDocument22 pagesForebrain Engraftment by Human Glial Progenitor Cells Enhances Synaptic Plasticity and Learning in Adult MiceAylin Lidsay Feria GPas encore d'évaluation
- (Advances in Cell Aging and Gerontology 17) Mark P. Mattson (Eds.) - Sleep and Aging-Academic Press, Elsevier (2005)Document192 pages(Advances in Cell Aging and Gerontology 17) Mark P. Mattson (Eds.) - Sleep and Aging-Academic Press, Elsevier (2005)Marcos R Galvão BatistaPas encore d'évaluation
- Neonate Movement Is Synchronized With Adult SpeechDocument3 pagesNeonate Movement Is Synchronized With Adult SpeechMarie Christine laznikPas encore d'évaluation
- Anafi Kayser and Raizen Nature Reviews Neuroscience 2018Document8 pagesAnafi Kayser and Raizen Nature Reviews Neuroscience 2018Francisco MtzPas encore d'évaluation
- Spinal Interneurons and Preganglionic Neurons in Sacral Autonomic Reflex PathwaysDocument15 pagesSpinal Interneurons and Preganglionic Neurons in Sacral Autonomic Reflex PathwaysNurRahMatPas encore d'évaluation
- The Neurologyof Sleep 2012 Published FormDocument18 pagesThe Neurologyof Sleep 2012 Published FormNeurologie PediatricaPas encore d'évaluation
- Communication From The Cerebellum To The Neocortex During Sleep SpindlesDocument1 pageCommunication From The Cerebellum To The Neocortex During Sleep SpindlesDavid NazarovPas encore d'évaluation
- Nihms 710720Document32 pagesNihms 710720Estigma Universidad Del RosarioPas encore d'évaluation
- Henn FA, Biol Psych 56, 2004Document5 pagesHenn FA, Biol Psych 56, 2004etaylorbensonPas encore d'évaluation
- Edwards - Neck Muscle Afferents Influence Oromotor and Cardiorespiratory CircuitsDocument16 pagesEdwards - Neck Muscle Afferents Influence Oromotor and Cardiorespiratory CircuitssbachorickPas encore d'évaluation
- UC San Diego Previously Published WorksDocument4 pagesUC San Diego Previously Published WorksGokushimakPas encore d'évaluation
- 12 ReferencesDocument26 pages12 ReferencesramopavelPas encore d'évaluation
- The Glymphatic System: A Novel Component of Fundamental NeurobiologyDocument14 pagesThe Glymphatic System: A Novel Component of Fundamental NeurobiologyramopavelPas encore d'évaluation
- Appendix Rotations in 3-D: A.1 Coordinate FramesDocument5 pagesAppendix Rotations in 3-D: A.1 Coordinate FramesramopavelPas encore d'évaluation
- Chapter Six Experimental Protocol: Study 1A 1B 2A 2B Subjects Stimulus TargetDocument15 pagesChapter Six Experimental Protocol: Study 1A 1B 2A 2B Subjects Stimulus TargetramopavelPas encore d'évaluation
- Chapter Two Ocular Motor System: Class Main Function Fixations Vestibular OptokineticDocument17 pagesChapter Two Ocular Motor System: Class Main Function Fixations Vestibular OptokineticramopavelPas encore d'évaluation
- Chapter Five Data Acquisition: 5.2a Search CoilsDocument7 pagesChapter Five Data Acquisition: 5.2a Search CoilsramopavelPas encore d'évaluation
- Aerobic Exerc Myotrophiesvoet2010Document34 pagesAerobic Exerc Myotrophiesvoet2010ramopavelPas encore d'évaluation
- Herbert Marcuse - Aggressiveness in Advanced Industrial SocietyDocument17 pagesHerbert Marcuse - Aggressiveness in Advanced Industrial SocietyHagay BarPas encore d'évaluation
- Chapter Eight Study OneDocument30 pagesChapter Eight Study OneramopavelPas encore d'évaluation
- Of Immune-Neuroendocrine Interactions: NetworkDocument12 pagesOf Immune-Neuroendocrine Interactions: NetworkramopavelPas encore d'évaluation
- Hell and Heaven, Nature and PersonDocument24 pagesHell and Heaven, Nature and PersonAlex ZammitPas encore d'évaluation
- Bosanquet Etal DMCN 2013Document9 pagesBosanquet Etal DMCN 2013ramopavelPas encore d'évaluation
- Chapter Three Vestibular SystemDocument22 pagesChapter Three Vestibular SystemramopavelPas encore d'évaluation
- MERS Coronavirus: Diagnostics, Epidemiology and TransmissionDocument21 pagesMERS Coronavirus: Diagnostics, Epidemiology and TransmissionramopavelPas encore d'évaluation
- Vestibulr NP Jn.00009.2016Document9 pagesVestibulr NP Jn.00009.2016ramopavelPas encore d'évaluation
- Loadin Startle ResponsDocument5 pagesLoadin Startle ResponsramopavelPas encore d'évaluation
- Journal of Clinical Microbiology-2020-Chan-JCM.00310-20.full PDFDocument33 pagesJournal of Clinical Microbiology-2020-Chan-JCM.00310-20.full PDFDwi Ratna KarimPas encore d'évaluation
- Hess Vest Phys Physiologyonline.2001.16.5.234Document5 pagesHess Vest Phys Physiologyonline.2001.16.5.234ramopavelPas encore d'évaluation
- Hess Vest Phys Physiologyonline.2001.16.5.234Document17 pagesHess Vest Phys Physiologyonline.2001.16.5.234ramopavelPas encore d'évaluation
- Escape From Homeostasis: Spinal Microcircuits and Progression of Amyotrophic Lateral SclerosisDocument13 pagesEscape From Homeostasis: Spinal Microcircuits and Progression of Amyotrophic Lateral SclerosisramopavelPas encore d'évaluation
- Einspieler Etal JPsychol 2008Document7 pagesEinspieler Etal JPsychol 2008ramopavelPas encore d'évaluation
- Alexandre Dumas 1845 Familia DeMediciDocument7 pagesAlexandre Dumas 1845 Familia DeMediciramopavelPas encore d'évaluation
- Vestibular Jn.01049.2015Document16 pagesVestibular Jn.01049.2015ramopavelPas encore d'évaluation
- Development of Postural Adjustments During Reaching in Infants at Risk For Cerebral Palsy From 4 To 18 MonthsDocument9 pagesDevelopment of Postural Adjustments During Reaching in Infants at Risk For Cerebral Palsy From 4 To 18 MonthsramopavelPas encore d'évaluation
- Alexandre Dumas 1845 Familia DeMediciDocument7 pagesAlexandre Dumas 1845 Familia DeMediciramopavelPas encore d'évaluation
- Vest jn.00078.2014Document11 pagesVest jn.00078.2014ramopavelPas encore d'évaluation
- Hess Vest Phys Physiologyonline.2001.16.5.234Document5 pagesHess Vest Phys Physiologyonline.2001.16.5.234ramopavelPas encore d'évaluation
- Konrad Lorenz The Analogy As A Source of KnowledgeDocument17 pagesKonrad Lorenz The Analogy As A Source of KnowledgeramopavelPas encore d'évaluation
- Cervical SpineDocument69 pagesCervical Spineveravero100% (3)
- Sci10 LM U3Document131 pagesSci10 LM U3Romel B Agno86% (7)
- Cns PathologyDocument18 pagesCns Pathologysunnyorange8Pas encore d'évaluation
- Assessing Sciatic Nerve Glide, Part II (Myofascial Techniques)Document4 pagesAssessing Sciatic Nerve Glide, Part II (Myofascial Techniques)Advanced-Trainings.com100% (7)
- All Type Essays - Biology emDocument27 pagesAll Type Essays - Biology emjoker boy100% (1)
- Human Nervous SystemDocument28 pagesHuman Nervous SystemMuhammad HaqPas encore d'évaluation
- Unifying Themes in The Study of LifeDocument7 pagesUnifying Themes in The Study of LifeKyla Renz de Leon100% (5)
- Chapter 17 Biology 2nd Year - Prof. Ijaz Ahmed Khan Abbasi (Lecturer Biology PGC) Notes - MDCAT by FUTURE DOCTORS - Touseef Ahmad Khan - 03499815886Document36 pagesChapter 17 Biology 2nd Year - Prof. Ijaz Ahmed Khan Abbasi (Lecturer Biology PGC) Notes - MDCAT by FUTURE DOCTORS - Touseef Ahmad Khan - 03499815886Suffa Academy100% (3)
- Geoffrey Jefferson (Contra Turing) - The Mind of Mechanical Man (CFR J Bruner)Document6 pagesGeoffrey Jefferson (Contra Turing) - The Mind of Mechanical Man (CFR J Bruner)Anonymous JIc7R6Pas encore d'évaluation
- Which of The Following Is The Component of Central Nervous System?Document28 pagesWhich of The Following Is The Component of Central Nervous System?nisaPas encore d'évaluation
- ALL DR Robert O. Becker: Silver Iontophoresis (Regeneration by Silver Ions)Document28 pagesALL DR Robert O. Becker: Silver Iontophoresis (Regeneration by Silver Ions)StellaEstel100% (1)
- Hes 029 Lab Sas 2Document2 pagesHes 029 Lab Sas 2Crislyn Jilou Bugas AdlawanPas encore d'évaluation
- The Main Parts of The Eye and Their FunctionDocument5 pagesThe Main Parts of The Eye and Their FunctionJoshuaPas encore d'évaluation
- Marcus Jacobson (Auth.) - Foundations of Neuroscience (1993)Document394 pagesMarcus Jacobson (Auth.) - Foundations of Neuroscience (1993)patata100% (1)
- 2301-Human Brain Apparent BoonDocument103 pages2301-Human Brain Apparent BoonSubramanya Seshagiri0% (1)
- CH 8 Nervous SystemPractice Test and KeyDocument13 pagesCH 8 Nervous SystemPractice Test and KeyvpcpharmdPas encore d'évaluation
- A. Multiple Choice. Answer TheDocument1 pageA. Multiple Choice. Answer TheJoeric CarinanPas encore d'évaluation
- Tissues - McqsDocument5 pagesTissues - McqsDocumentSharerPas encore d'évaluation
- PDF - Sat Practice Test 10 Answers PDFDocument50 pagesPDF - Sat Practice Test 10 Answers PDFAryan ShahPas encore d'évaluation
- The MELT Method by Sue HitzmannDocument20 pagesThe MELT Method by Sue HitzmannHarperOne (an imprint of HarperCollins)67% (6)
- Definition of Medical EtymologyDocument2 pagesDefinition of Medical EtymologyOny Hertriandita APas encore d'évaluation
- 06a - Introduction To Spinal Cord - NeuroanatomyDocument9 pages06a - Introduction To Spinal Cord - Neuroanatomyhiba jasimPas encore d'évaluation
- Science6 q2 Mod3 Explain How The Different Organ System v2Document28 pagesScience6 q2 Mod3 Explain How The Different Organ System v2Mark James VallejosPas encore d'évaluation
- Neurotoxin Protocol Jan06Document19 pagesNeurotoxin Protocol Jan06mary eng100% (2)
- Sistem SarafDocument141 pagesSistem SarafPei YiingPas encore d'évaluation
- Brain NeuroanatomyDocument227 pagesBrain NeuroanatomyNkechukwu OgakwuPas encore d'évaluation
- SUSAN R - JOHNSON, M - D - On Television and Our Children's MindsDocument11 pagesSUSAN R - JOHNSON, M - D - On Television and Our Children's MindsSusan HoekstraPas encore d'évaluation
- 2014 Neurogenic NeuroinflammationDocument11 pages2014 Neurogenic NeuroinflammationrendyjiwonoPas encore d'évaluation
- Neural Mobilizationw10Document33 pagesNeural Mobilizationw10manjukumard2007Pas encore d'évaluation
- Meningitis (Deadly Diseases and Epidemics) (PDFDrive)Document121 pagesMeningitis (Deadly Diseases and Epidemics) (PDFDrive)Islamiyatul Hayati MiswanPas encore d'évaluation
- Biological Basis of PersonalityDocument58 pagesBiological Basis of PersonalityAldrian Maulion EvangelistaPas encore d'évaluation