Vous aimerez peut-être aussi
- BienvenidaDocument3 pagesBienvenidaNatsumiTakamoriPas encore d'évaluation
- Contenido Tematico Ii Bimestre - Biología - 3secDocument1 pageContenido Tematico Ii Bimestre - Biología - 3secNatsumiTakamoriPas encore d'évaluation
- s1 Recuperacion Secundaria4 Experiencias 1Document20 pagess1 Recuperacion Secundaria4 Experiencias 1Angela Apaza Torres0% (1)
- Taller de CienciasDocument4 pagesTaller de CienciasNatsumiTakamoriPas encore d'évaluation
- Contenido Tematico Ii Bimestre - Biología - 1secDocument1 pageContenido Tematico Ii Bimestre - Biología - 1secNatsumiTakamoriPas encore d'évaluation
- Elaboración de EncurtidosDocument4 pagesElaboración de EncurtidosNatsumiTakamori100% (2)
- MÚSCULOSDocument27 pagesMÚSCULOSNatsumiTakamoriPas encore d'évaluation
- Contenido Tematico Ii Bimestre - Biología - 5secDocument1 pageContenido Tematico Ii Bimestre - Biología - 5secNatsumiTakamoriPas encore d'évaluation
- Contenido Tematico Ii Bimestre - Biología - 4secDocument1 pageContenido Tematico Ii Bimestre - Biología - 4secNatsumiTakamoriPas encore d'évaluation
- Directorio OdpeDocument3 pagesDirectorio OdpeRocioRamosJaraPas encore d'évaluation
- ViroDocument9 pagesViroNatsumiTakamoriPas encore d'évaluation
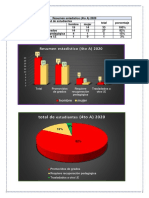
- Resumen Estadístico 5 SecDocument3 pagesResumen Estadístico 5 SecNatsumiTakamoriPas encore d'évaluation
- RevisiónDocument20 pagesRevisiónNatsumiTakamoriPas encore d'évaluation
- Frutos: morfología y clasificaciónDocument4 pagesFrutos: morfología y clasificaciónNatsumiTakamoriPas encore d'évaluation
- 1 BMEnzimas Biología Molecular 2018Document55 pages1 BMEnzimas Biología Molecular 2018NatsumiTakamoriPas encore d'évaluation
- Deshidratación y rehidratación en peces, anfibios y crustáceosDocument3 pagesDeshidratación y rehidratación en peces, anfibios y crustáceosNatsumiTakamoriPas encore d'évaluation
- PedigriDocument8 pagesPedigriNatsumiTakamoriPas encore d'évaluation
- Semana 15 MEDIO AMBIENTE TÉRMICODocument8 pagesSemana 15 MEDIO AMBIENTE TÉRMICONatsumiTakamoriPas encore d'évaluation
- Lectura 9 Anomalías NuméricasDocument8 pagesLectura 9 Anomalías NuméricasLeslie Vanessa Vargas BernuyPas encore d'évaluation
- Paper Proteinas de Union A DNADocument10 pagesPaper Proteinas de Union A DNASebastian GodoyPas encore d'évaluation
- BiotecnologiaDocument7 pagesBiotecnologiaNatsumiTakamoriPas encore d'évaluation
- 22mutaciones PDFDocument10 pages22mutaciones PDFDiana GonzálezPas encore d'évaluation
- Estudio de la variación espacio-temporal de la comunidad vegetal de las Lomas de CarabaylloDocument140 pagesEstudio de la variación espacio-temporal de la comunidad vegetal de las Lomas de CarabaylloNatsumiTakamoriPas encore d'évaluation
- Capítulo 5Document2 pagesCapítulo 5NatsumiTakamoriPas encore d'évaluation
- Art 10ZA12 PDFDocument18 pagesArt 10ZA12 PDFNatsumiTakamoriPas encore d'évaluation
- Técnicas inmunológicas secundariasDocument4 pagesTécnicas inmunológicas secundariasdarkzero_858Pas encore d'évaluation
- BiotecnologiaDocument7 pagesBiotecnologiaNatsumiTakamoriPas encore d'évaluation
- Laboratorio N8 Identificacion de Cromatina Sexual PDFDocument3 pagesLaboratorio N8 Identificacion de Cromatina Sexual PDFNatsumiTakamoriPas encore d'évaluation
- 1 PBDocument3 pages1 PBNatsumiTakamoriPas encore d'évaluation
- Las Lomas de Lima - MamaniDocument25 pagesLas Lomas de Lima - MamaniIván D. Delgado PugleyPas encore d'évaluation
- Ejeraut 1Document5 pagesEjeraut 1pepepepePas encore d'évaluation
- Efecto Del Nitrato, Irradiancia y Salinidad Sobre La Producción de Clorofila A Partir de Microalgas Aisladas en La Región Noroccidental de VenezuelaDocument11 pagesEfecto Del Nitrato, Irradiancia y Salinidad Sobre La Producción de Clorofila A Partir de Microalgas Aisladas en La Región Noroccidental de VenezuelajoselbermudezgPas encore d'évaluation
- Nom 006 Asea 2017 FinalDocument114 pagesNom 006 Asea 2017 FinalJAVIER VALENCIAPas encore d'évaluation
- Estructura de Las Proteínas 12Document44 pagesEstructura de Las Proteínas 12uliszerPas encore d'évaluation
- Diseño ExperimentalDocument6 pagesDiseño Experimentalronal muñozPas encore d'évaluation
- Informe de Obtencion de HidrocarburosDocument9 pagesInforme de Obtencion de HidrocarburosAdrian VelascoPas encore d'évaluation
- Espectroscopía EjemplosDocument14 pagesEspectroscopía EjemplosSule Granados DPas encore d'évaluation
- HistorialDocument27 pagesHistorialTaty Samaniego NavarroPas encore d'évaluation
- Triptico de Seguridad en El LaboratorioDocument1 pageTriptico de Seguridad en El Laboratoriodepablo08Pas encore d'évaluation
- Clasificación de los aminoácidos según sus propiedadesDocument2 pagesClasificación de los aminoácidos según sus propiedadesJosmel yañezPas encore d'évaluation
- Cinética EnzimáticaDocument36 pagesCinética EnzimáticaYaneth AguilarPas encore d'évaluation
- Nomenclatura inorgánica: prueba parcial de químicaDocument5 pagesNomenclatura inorgánica: prueba parcial de químicaNataly Neira GuzmánPas encore d'évaluation
- Unidad 2 Enlaces Quimicos - Clasificacion de Enlaces QuimicosDocument29 pagesUnidad 2 Enlaces Quimicos - Clasificacion de Enlaces QuimicosMarcos SolisPas encore d'évaluation
- ACEITE Chichipe Lozano Jhon Percy ACEITE DE TAMARINDODocument47 pagesACEITE Chichipe Lozano Jhon Percy ACEITE DE TAMARINDOAdriano Ortega SánchezPas encore d'évaluation
- Factores Que Influyen en La Toxicidad de Una SustnciaDocument10 pagesFactores Que Influyen en La Toxicidad de Una SustnciaIng Max de la CruzPas encore d'évaluation
- Mapa Conceptual BiologiaDocument1 pageMapa Conceptual BiologiaCatalina Belén CornejoPas encore d'évaluation
- Guía Tema IIDocument23 pagesGuía Tema IIkariPas encore d'évaluation
- Guia 2P. Química III. 2022.Document2 pagesGuia 2P. Química III. 2022.Ana AguilarPas encore d'évaluation
- SAPONIFICACIÓNDocument6 pagesSAPONIFICACIÓNjoel_garcia_112Pas encore d'évaluation
- Práctica 3 EmbutidosDocument10 pagesPráctica 3 EmbutidosRaquel LaraPas encore d'évaluation
- VIKI VEREAU VADEMECUM 2016 RW - 12.01.16Document100 pagesVIKI VEREAU VADEMECUM 2016 RW - 12.01.16Irina Kucevichute100% (1)
- Informe SolubilidadDocument8 pagesInforme SolubilidadZoila Jimenez JimenezPas encore d'évaluation
- Determinacion de AcidezDocument3 pagesDeterminacion de AcidezdayanaPas encore d'évaluation
- Sintesis de CumarimaDocument5 pagesSintesis de CumarimammaarrPas encore d'évaluation
- SÍNTESIS AMARILLO MARTIUS SUSTITUCIÓN ELECTROFÍLICA 1-NAFTOLDocument5 pagesSÍNTESIS AMARILLO MARTIUS SUSTITUCIÓN ELECTROFÍLICA 1-NAFTOLDiaz AndresPas encore d'évaluation
- Ejercicios Propuestos de Hibridación Del CarbonoDocument4 pagesEjercicios Propuestos de Hibridación Del CarbonoKarlitah OjedaPas encore d'évaluation
- Amidas y Aminas 10mo DDocument17 pagesAmidas y Aminas 10mo DEnrique Rojas BermudoPas encore d'évaluation
- CatalogoMAYJUN2023WHATSAPP 1686950433109Document112 pagesCatalogoMAYJUN2023WHATSAPP 1686950433109Maria Isabel Naranjo HincapiePas encore d'évaluation
- Metabolismo Celular Bio LabDocument6 pagesMetabolismo Celular Bio LabAngello Cristobal VictorioPas encore d'évaluation
- Jabones y DetergentesDocument8 pagesJabones y DetergentesJosseline ObregónPas encore d'évaluation