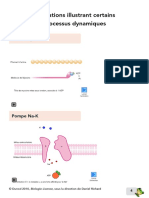
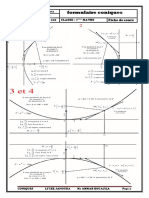
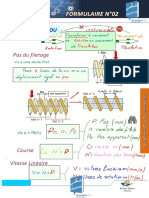
Vous aimerez peut-être aussi
- COURS - 3 - Procaryote EucaryoteDocument40 pagesCOURS - 3 - Procaryote EucaryoteNESRINE NABIPas encore d'évaluation
- Chapitre I Structure de La CelluleDocument107 pagesChapitre I Structure de La CelluleDescartes KpannePas encore d'évaluation
- La Cellule EucaryoteDocument9 pagesLa Cellule Eucaryoteherrere64Pas encore d'évaluation
- Cours 1 - Introduction À La Biologie CellulaireDocument30 pagesCours 1 - Introduction À La Biologie Cellulaireamina44Pas encore d'évaluation
- Membranne Plasmique CompressedDocument81 pagesMembranne Plasmique CompressedRedouan KamalyPas encore d'évaluation
- Procréation: Les Grands Articles d'UniversalisD'EverandProcréation: Les Grands Articles d'UniversalisPas encore d'évaluation
- Photosynthèse: Les Grands Articles d'UniversalisD'EverandPhotosynthèse: Les Grands Articles d'UniversalisPas encore d'évaluation
- Développement végétal: Les Grands Articles d'UniversalisD'EverandDéveloppement végétal: Les Grands Articles d'UniversalisPas encore d'évaluation
- Génétique: Les Grands Articles d'UniversalisD'EverandGénétique: Les Grands Articles d'UniversalisPas encore d'évaluation
- Cours BioCell Chap1et2Document107 pagesCours BioCell Chap1et2NancyPas encore d'évaluation
- Snv-Biologie Cellulaire-Les Membranes CellulairesDocument5 pagesSnv-Biologie Cellulaire-Les Membranes Cellulairesamira messoudPas encore d'évaluation
- Unité 1 - La Cellule, Unité Du VivantDocument8 pagesUnité 1 - La Cellule, Unité Du VivantIrene PGPas encore d'évaluation
- COURS Enérgétique 2017Document15 pagesCOURS Enérgétique 2017ChaiMaePas encore d'évaluation
- La CelluleDocument6 pagesLa CelluleHossam AnaPas encore d'évaluation
- Biologie Cellulaire Et Biochimie Structural 2Document14 pagesBiologie Cellulaire Et Biochimie Structural 2bastaouiiPas encore d'évaluation
- COURS BCF EtudiantsDocument22 pagesCOURS BCF Etudiantsamine bedjaPas encore d'évaluation
- Biologie CellulaireDocument24 pagesBiologie CellulairealmnaouarPas encore d'évaluation
- Cours 1 Culture CellulaireDocument9 pagesCours 1 Culture CellulaireLabgaa SarahPas encore d'évaluation
- BIO 111 - CHAP - III - IV - Et - VDocument8 pagesBIO 111 - CHAP - III - IV - Et - VBerthe Aristide Ngue NkongoPas encore d'évaluation
- Biologie Chapitre1Document30 pagesBiologie Chapitre1Nada Tahiri100% (1)
- Cellules Procaryotes Et Cellules EucaryotesDocument10 pagesCellules Procaryotes Et Cellules EucaryotesalmnaouarPas encore d'évaluation
- Cours SystematiqueDocument99 pagesCours SystematiquemokermiPas encore d'évaluation
- Fma1s1-Biochimie Structurale PR Balouch V-08092018Document105 pagesFma1s1-Biochimie Structurale PR Balouch V-08092018Féz EyPas encore d'évaluation
- Cours Complet de BiochimieDocument124 pagesCours Complet de BiochimieNiamatelah LahkimPas encore d'évaluation
- Physiologie Vegetale 2 S4Document20 pagesPhysiologie Vegetale 2 S4hassan aitPas encore d'évaluation
- Biocell 1 - Cytologie (Juin 2019, Laura T)Document220 pagesBiocell 1 - Cytologie (Juin 2019, Laura T)Daniela RiosPas encore d'évaluation
- SalutDocument1 pageSalutSalah eddinePas encore d'évaluation
- CytosqueletteDocument24 pagesCytosquelettefellah aminaPas encore d'évaluation
- Biologie Des Acides NucléiquesDocument22 pagesBiologie Des Acides Nucléiquesabekhti2008Pas encore d'évaluation
- Polycopie Cours Microbiologie Imziln 2015 16Document34 pagesPolycopie Cours Microbiologie Imziln 2015 16yassin assebanPas encore d'évaluation
- Chapitre 3 La Cellule - Les OrganitesDocument43 pagesChapitre 3 La Cellule - Les Organitesahmed daoudi100% (1)
- TD Et Corrigés - Ecologie Végétale - SV3 2018 PDFDocument26 pagesTD Et Corrigés - Ecologie Végétale - SV3 2018 PDFDJIL BENPas encore d'évaluation
- TD Enzymo Biochimie Met 3Document5 pagesTD Enzymo Biochimie Met 3Ali SabPas encore d'évaluation
- L'asymétrie Des Membranes BiologiquesDocument14 pagesL'asymétrie Des Membranes Biologiquessafiadjili100% (1)
- Cours de Biologie Végétale Pour L1 BOUZID SalhaDocument82 pagesCours de Biologie Végétale Pour L1 BOUZID SalhaMahamadi Belem100% (1)
- Structure Et Anatomie Fonctionnelle Des Bacteries-ConvertiDocument55 pagesStructure Et Anatomie Fonctionnelle Des Bacteries-ConvertiAmiraBenhammou100% (1)
- Bio Cell Chap 1+2 + 3Document49 pagesBio Cell Chap 1+2 + 3ManarPas encore d'évaluation
- Ogm Et Mutations Sabine MehoumeDocument15 pagesOgm Et Mutations Sabine MehoumeMarius Ganhouegnon100% (1)
- 2100534491biologie PDFDocument712 pages2100534491biologie PDFaloys Ndzie100% (5)
- Chapitre 2 - La Production de Matière Par Les Cellules ChlorophylliennesDocument6 pagesChapitre 2 - La Production de Matière Par Les Cellules ChlorophylliennesjoanaPas encore d'évaluation
- Biologie CellulaireDocument24 pagesBiologie Cellulairealmnaouar100% (2)
- Ts Genetique RemerandDocument33 pagesTs Genetique Remerandbeebac2009Pas encore d'évaluation
- Bio CellulaireDocument44 pagesBio CellulaireTouri Na100% (1)
- Microbiologie S3Document87 pagesMicrobiologie S3api-3738916100% (9)
- (PACES) Physiologie Humaine by Hervé GuénardDocument1 371 pages(PACES) Physiologie Humaine by Hervé Guénardsam egoroffPas encore d'évaluation
- M Triqui Cours de Biologie Cellulaire 2016-2017 PDFDocument105 pagesM Triqui Cours de Biologie Cellulaire 2016-2017 PDFRami Nadjm80% (5)
- Compl Biologie Etudiant S-1Document43 pagesCompl Biologie Etudiant S-1aloys NdziePas encore d'évaluation
- Biologie VegetaleDocument16 pagesBiologie Vegetalekhawla amamouPas encore d'évaluation
- BiochimieDocument78 pagesBiochimieZoumana CoulibalyPas encore d'évaluation
- Biophysique Cours 1Document105 pagesBiophysique Cours 1Smail FreeManPas encore d'évaluation
- Chap1 EucaryoteDocument4 pagesChap1 Eucaryoteslimdamak100% (1)
- L Essentiel de La Biologie Cellulaire 3 Ed Chapitre16Document18 pagesL Essentiel de La Biologie Cellulaire 3 Ed Chapitre16Vayo SonyPas encore d'évaluation
- Chapitre I BiologieDocument26 pagesChapitre I BiologieDjedje DjellabPas encore d'évaluation
- La Cellule 1Document29 pagesLa Cellule 1ۥٰۥٰۥٰ ۥٰۥٰۥٰPas encore d'évaluation
- 1er Cours Introduction Au Monde MicrobienDocument8 pages1er Cours Introduction Au Monde MicrobienMoufida Trénitarya100% (1)
- Cours Lipides - Part 1Document74 pagesCours Lipides - Part 1AhmedPas encore d'évaluation
- Chap 01Document33 pagesChap 01hamidaPas encore d'évaluation
- La PhotosyntheseDocument24 pagesLa Photosynthesetoto TOTOROTOPas encore d'évaluation
- BIO 270 Physiologie Cellulaire Générale 2020 PDFDocument125 pagesBIO 270 Physiologie Cellulaire Générale 2020 PDFTONDI ABDOU CHACOURPas encore d'évaluation
- Chapitre 4 + 5 Cycle Cellulaire Et Noyau InterphasiqueDocument15 pagesChapitre 4 + 5 Cycle Cellulaire Et Noyau InterphasiqueOuissem Belkhous100% (1)
- Tri&Recherche PDFDocument4 pagesTri&Recherche PDFBelgasem AsselPas encore d'évaluation
- Serie RécursivitéDocument3 pagesSerie RécursivitéBelgasem Assel100% (1)
- Cours SQL - Base de DonnéesDocument30 pagesCours SQL - Base de DonnéesRouaissi RidhaPas encore d'évaluation
- Catalyse L3Document85 pagesCatalyse L3Wassi La100% (1)
- 3 Exos Genetique2 PDFDocument3 pages3 Exos Genetique2 PDFLotfi Touil100% (1)
- Exercices Corrig S - G N Tique HumaineDocument7 pagesExercices Corrig S - G N Tique HumaineBelgasem Assel100% (1)
- Correction Continuité Limites 4ème Mathématiques1 PDFDocument7 pagesCorrection Continuité Limites 4ème Mathématiques1 PDFBelgasem AsselPas encore d'évaluation
- 25 Reac ChiDocument23 pages25 Reac ChiBelgasem AsselPas encore d'évaluation
- Base de Données: I. Introduction GénéraleDocument13 pagesBase de Données: I. Introduction GénéraleBelgasem AsselPas encore d'évaluation
- 4M Serie2 - Parabole 2020Document3 pages4M Serie2 - Parabole 2020Belgasem AsselPas encore d'évaluation
- 4M Serie1 - Parabole 2020 CorDocument4 pages4M Serie1 - Parabole 2020 CorBelgasem AsselPas encore d'évaluation
- 2eme Seance 18 Sept2017 Diagrammes Unaires Suite Introduction Diagrammes BinairesDocument32 pages2eme Seance 18 Sept2017 Diagrammes Unaires Suite Introduction Diagrammes BinairesBelgasem AsselPas encore d'évaluation
- Demcours 02Document1 pageDemcours 02Belgasem AsselPas encore d'évaluation
- Ex 20 CorDocument3 pagesEx 20 CorBelgasem AsselPas encore d'évaluation
- DSN 1Document8 pagesDSN 1Belgasem AsselPas encore d'évaluation
- Ex 15Document1 pageEx 15Belgasem AsselPas encore d'évaluation
- 4.i.revision - Gueddiche.zoubaier 08.09.cor PDFDocument5 pages4.i.revision - Gueddiche.zoubaier 08.09.cor PDFBelgasem AsselPas encore d'évaluation
- La Fonction Reproductrice FeminineDocument2 pagesLa Fonction Reproductrice FeminineBelgasem AsselPas encore d'évaluation
- Demcours 04Document1 pageDemcours 04Belgasem AsselPas encore d'évaluation
- Demcours 01Document1 pageDemcours 01Belgasem AsselPas encore d'évaluation
- Ex 01Document1 pageEx 01Belgasem AsselPas encore d'évaluation
- 2sex Int Lim Cont ExeDocument4 pages2sex Int Lim Cont ExebidaouimounirPas encore d'évaluation
- Devoir de Synthèse N°3 - Économie - Bac Eco (2010-2011) MR ZribiDocument2 pagesDevoir de Synthèse N°3 - Économie - Bac Eco (2010-2011) MR ZribiBelgasem AsselPas encore d'évaluation
- A463db PDFDocument3 pagesA463db PDFBelgasem AsselPas encore d'évaluation
- These Doctor atDocument47 pagesThese Doctor atBelgasem AsselPas encore d'évaluation
- Convertisseur WebDocument2 pagesConvertisseur WebBelgasem AsselPas encore d'évaluation
- Transfor de MouvmtDocument8 pagesTransfor de MouvmtBelgasem AsselPas encore d'évaluation
- ch12 1pilesDocument4 pagesch12 1pilesBelgasem AsselPas encore d'évaluation
- 2006 DessinDocument2 pages2006 DessinBelgasem AsselPas encore d'évaluation
- La BiochimieDocument44 pagesLa BiochimieDjawed BoutPas encore d'évaluation
- Les Étapes Du Séquençage d'ADN... - Biologie Université Facebook PDFDocument1 pageLes Étapes Du Séquençage d'ADN... - Biologie Université Facebook PDFAnas DiouriPas encore d'évaluation
- Ecologie MicrobienneDocument25 pagesEcologie MicrobiennesamPas encore d'évaluation
- Épinutrition - Denis RichéDocument97 pagesÉpinutrition - Denis RichéDjamel BaboriPas encore d'évaluation
- La Découverte de La Molécule ADN - CorrectionDocument3 pagesLa Découverte de La Molécule ADN - CorrectionBeshta AhmadiPas encore d'évaluation
- BTS Dietetique Les Acides NucleiquesDocument4 pagesBTS Dietetique Les Acides NucleiquesEkissi Etienne OrsotPas encore d'évaluation
- TD Microbio M1 Generalites Reponses PDFDocument11 pagesTD Microbio M1 Generalites Reponses PDFSaber Ben ZianPas encore d'évaluation
- Molecul2 Reparation ReplicationDocument7 pagesMolecul2 Reparation Replicationhappy nefPas encore d'évaluation
- 2as Benbali Tayeb1Document5 pages2as Benbali Tayeb1tayebPas encore d'évaluation
- Glucides Lipides Protéines Acides NucléiquesDocument41 pagesGlucides Lipides Protéines Acides NucléiquesAchwak BelfadelPas encore d'évaluation
- QCMDocument10 pagesQCMFethiSvtPas encore d'évaluation
- AaaaaaaaDocument36 pagesAaaaaaaamaysounPas encore d'évaluation
- Biologie VégétaleDocument84 pagesBiologie VégétaleIliass Naouadir100% (8)
- FR HSG Microbiology - V3 2022 MR 031Document1 pageFR HSG Microbiology - V3 2022 MR 031Sigma QualPas encore d'évaluation
- La Synthèse ProtéiqueDocument6 pagesLa Synthèse ProtéiquealiPas encore d'évaluation
- La Correction - L Expression Du Patrimoine Genetique 2Document8 pagesLa Correction - L Expression Du Patrimoine Genetique 2amal aliPas encore d'évaluation
- Examen de Biologie 2020Document3 pagesExamen de Biologie 2020LisaPas encore d'évaluation
- Replication de l'ADN (1) - 020827Document29 pagesReplication de l'ADN (1) - 020827nonamethe427Pas encore d'évaluation
- Manuel D'utilisation Amplix 12A FR V2Document40 pagesManuel D'utilisation Amplix 12A FR V2Abdelali EnnouariPas encore d'évaluation
- ImmunoDocument29 pagesImmunoJalelo AbdelPas encore d'évaluation
- Traitement de L'eau - Les Oxydants Et Désinfectants - Degremont®Document24 pagesTraitement de L'eau - Les Oxydants Et Désinfectants - Degremont®Chaoubi YoussefPas encore d'évaluation
- Cours Génétique LPSVT S3Document87 pagesCours Génétique LPSVT S3samifilol703Pas encore d'évaluation
- Biochimie - Pré-Rentrée S1 - Activité 1 MacromoléculesDocument112 pagesBiochimie - Pré-Rentrée S1 - Activité 1 MacromoléculesNathan CohenPas encore d'évaluation
- CODEX BIOGENESIS: THE Book On DNA Numerical Decoding...Document1 pageCODEX BIOGENESIS: THE Book On DNA Numerical Decoding...Jean-claude PerezPas encore d'évaluation
- ESVG Chap1 - DS 1 Exo Mitose Et BrdU CORR-2 PDFDocument3 pagesESVG Chap1 - DS 1 Exo Mitose Et BrdU CORR-2 PDFJamal Aboussaid100% (3)
- TD AdnDocument4 pagesTD AdnHABRIOUX FabiennePas encore d'évaluation
- Biotech Mutagenese PDFDocument8 pagesBiotech Mutagenese PDFAghiles Ardj100% (2)
- Les Conséquences de La Variabilité de l'ADNDocument8 pagesLes Conséquences de La Variabilité de l'ADNximena silvaPas encore d'évaluation
- Le Noyau SciencesDocument2 pagesLe Noyau SciencesGauthier MehaudensPas encore d'évaluation
- 6a p2 Ue2 Cours 7 Radiobiologie Radioprotection Ronéo 2Document14 pages6a p2 Ue2 Cours 7 Radiobiologie Radioprotection Ronéo 2Idrissa Mama DiarraPas encore d'évaluation