Vous aimerez peut-être aussi
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- Form, Function, Food and Feeding in Stellate Echinoderms : ZoosymposiaDocument10 pagesForm, Function, Food and Feeding in Stellate Echinoderms : ZoosymposiaSixto Gutiérrez SaavedraPas encore d'évaluation
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (587)
- PAP Arendt Et Al 2015 PTRSBDocument18 pagesPAP Arendt Et Al 2015 PTRSBSixto Gutiérrez SaavedraPas encore d'évaluation
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (894)
- Microbiota-Gut-Brain Axis Mechanisms in The Complex Network PDFDocument29 pagesMicrobiota-Gut-Brain Axis Mechanisms in The Complex Network PDFbenqtenPas encore d'évaluation
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- The Blood-Brain Barrier in Aging and NeurodegenerationDocument15 pagesThe Blood-Brain Barrier in Aging and NeurodegenerationBruno FagundesPas encore d'évaluation
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- PAP Apitz & Agius 2013 MPBDocument15 pagesPAP Apitz & Agius 2013 MPBSixto Gutiérrez SaavedraPas encore d'évaluation
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- Nature 09678Document7 pagesNature 09678Cesar Valenzuela EncinasPas encore d'évaluation
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- Eme 2017Document14 pagesEme 2017Sixto Gutiérrez SaavedraPas encore d'évaluation
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- PAP Dey Et Al 2016 Cell BiolDocument10 pagesPAP Dey Et Al 2016 Cell BiolSixto Gutiérrez SaavedraPas encore d'évaluation
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- News & Analysis: Reshaping The Tree of LifeDocument1 pageNews & Analysis: Reshaping The Tree of LifeSixto Gutiérrez SaavedraPas encore d'évaluation
- PAP Baena-Nogueras Et Al 2018 MPBDocument10 pagesPAP Baena-Nogueras Et Al 2018 MPBSixto Gutiérrez SaavedraPas encore d'évaluation
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- s41467 018 07214 WDocument13 pagess41467 018 07214 WSixto Gutiérrez SaavedraPas encore d'évaluation
- PAP Apitz Et Al 2017a IEAMDocument11 pagesPAP Apitz Et Al 2017a IEAMSixto Gutiérrez SaavedraPas encore d'évaluation
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Zhou 2018Document10 pagesZhou 2018Sixto Gutiérrez SaavedraPas encore d'évaluation
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- PAP Haro Et Al 2013 AIPDocument14 pagesPAP Haro Et Al 2013 AIPSixto Gutiérrez SaavedraPas encore d'évaluation
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2219)
- PAP Backhaus Et Al 2016 IEAMDocument1 pagePAP Backhaus Et Al 2016 IEAMSixto Gutiérrez SaavedraPas encore d'évaluation
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- PAP Assis Et Al 2017 GEBDocument8 pagesPAP Assis Et Al 2017 GEBSixto Gutiérrez SaavedraPas encore d'évaluation
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- PAP Abaei Et Al 2016 Renew EnergyDocument22 pagesPAP Abaei Et Al 2016 Renew EnergySixto Gutiérrez SaavedraPas encore d'évaluation
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (265)
- Pap Ali Et Al 2015 MjasDocument7 pagesPap Ali Et Al 2015 MjasSixto Gutiérrez SaavedraPas encore d'évaluation
- PAP - Apitz Et Al - 2017a - IEAM PDFDocument14 pagesPAP - Apitz Et Al - 2017a - IEAM PDFSixto Gutiérrez SaavedraPas encore d'évaluation
- CRU Iwasa-Arai & Serejo 2018 ZJLSDocument29 pagesCRU Iwasa-Arai & Serejo 2018 ZJLSSixto Gutiérrez SaavedraPas encore d'évaluation
- PAP Arblaster Et Al 2015 IEAMDocument12 pagesPAP Arblaster Et Al 2015 IEAMSixto Gutiérrez SaavedraPas encore d'évaluation
- The Revised Classification of EukaryotesDocument79 pagesThe Revised Classification of Eukaryoteskrishy19Pas encore d'évaluation
- Neff 2011Document37 pagesNeff 2011Sixto Gutiérrez SaavedraPas encore d'évaluation
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- PAP Caswell Et Al 2018 MPBDocument18 pagesPAP Caswell Et Al 2018 MPBSixto Gutiérrez SaavedraPas encore d'évaluation
- PAP Admed Et Al 2018 MPBDocument12 pagesPAP Admed Et Al 2018 MPBSixto Gutiérrez SaavedraPas encore d'évaluation
- Fisher 2017Document27 pagesFisher 2017Sixto Gutiérrez SaavedraPas encore d'évaluation
- Chemosphere: Stuart L. Simpson, David A. Spadaro, Dom O'BrienDocument8 pagesChemosphere: Stuart L. Simpson, David A. Spadaro, Dom O'BrienSixto Gutiérrez SaavedraPas encore d'évaluation
- Assessment of Environmental Quality of Marine Sediments in The Bay System Caldera (27° S), ChileDocument18 pagesAssessment of Environmental Quality of Marine Sediments in The Bay System Caldera (27° S), ChileSixto Gutiérrez SaavedraPas encore d'évaluation
- Bay 2012Document13 pagesBay 2012Sixto Gutiérrez SaavedraPas encore d'évaluation
- Human Genome ProjectDocument13 pagesHuman Genome Projectbszool006Pas encore d'évaluation
- Effects of Air Pollutants On Mediterranean and Temperate Forest EcosystemsDocument263 pagesEffects of Air Pollutants On Mediterranean and Temperate Forest EcosystemsPACIFIC SOUTHWEST RESEARCH STATION REPORTPas encore d'évaluation
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (119)
- Chapter 15 - TeratogenesisDocument29 pagesChapter 15 - TeratogenesisMaula AhmadPas encore d'évaluation
- To Study The Quantity Case in Present in Different Sample of MilkDocument7 pagesTo Study The Quantity Case in Present in Different Sample of Milkbelly4u100% (2)
- Bio-Diversity Uses Threats and ConservationDocument32 pagesBio-Diversity Uses Threats and ConservationRoshin VarghesePas encore d'évaluation
- Campion Mumford Morgeson Nahrgang 2005 PDFDocument24 pagesCampion Mumford Morgeson Nahrgang 2005 PDFioanaelena89Pas encore d'évaluation
- MSC QuizDocument6 pagesMSC QuizLokesh WaranPas encore d'évaluation
- Jhean Steffan Martines de CamargoDocument2 pagesJhean Steffan Martines de CamargoJheanPas encore d'évaluation
- Fish Reproduction StrategiesDocument30 pagesFish Reproduction StrategiesWahyuniFanggiTasikPas encore d'évaluation
- Cell Structure and Function ChapterDocument80 pagesCell Structure and Function ChapterDrAmit Verma0% (2)
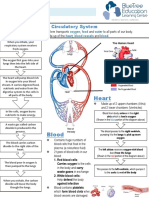
- Circulatory System Notes SLDocument1 pageCirculatory System Notes SLRathna PalaniappanPas encore d'évaluation
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Mcws Pamp (Final Pamp - Aug 8 2016)Document102 pagesMcws Pamp (Final Pamp - Aug 8 2016)Mount Calavite DENRPas encore d'évaluation
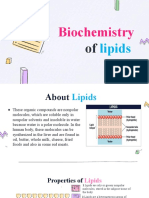
- Lipid PresentationDocument20 pagesLipid PresentationFrances Anne OrbinoPas encore d'évaluation
- Study Guide Lecture Exam 1 Use This Handout With Your Video Review Possible Essay QuestionsDocument4 pagesStudy Guide Lecture Exam 1 Use This Handout With Your Video Review Possible Essay QuestionsBrandice BradleyPas encore d'évaluation
- Notes - Growth and DevelopmentDocument4 pagesNotes - Growth and DevelopmentNathan mwapePas encore d'évaluation
- Birds of Prey PDFDocument24 pagesBirds of Prey PDFPaula MatosPas encore d'évaluation
- SouthPath Reference RangesDocument1 pageSouthPath Reference Rangesddp2710Pas encore d'évaluation
- Comprehension QuestionDocument2 pagesComprehension QuestionAhmad Safuan Bin Mohd Sukri -Pas encore d'évaluation
- 5.2 BiophiliaDocument24 pages5.2 BiophiliaNurFarahinZakariahPas encore d'évaluation
- Udc Vol 1Document921 pagesUdc Vol 1Dani TrashPas encore d'évaluation
- Soal Pat Inggris Kelas 7 - 2021Document9 pagesSoal Pat Inggris Kelas 7 - 2021Bella Septiani FaryanPas encore d'évaluation
- The Heart: Structure and FunctionDocument2 pagesThe Heart: Structure and FunctionHolly May MontejoPas encore d'évaluation
- Organizational Behavior Field Map and Perception FactorsDocument32 pagesOrganizational Behavior Field Map and Perception FactorsPratik TagwalePas encore d'évaluation
- Wbtet 2012Document7 pagesWbtet 2012Partha BatabyalPas encore d'évaluation
- Types of COVID VaccinesDocument1 pageTypes of COVID VaccinesEmre ÇELİKPas encore d'évaluation
- Synthesis and Characterization of EDTA ComplexesDocument7 pagesSynthesis and Characterization of EDTA ComplexesLucas PereiraPas encore d'évaluation
- Dr. Morse's Herbal Formulation ListDocument10 pagesDr. Morse's Herbal Formulation ListTom50% (2)
- Advanced Powder Technology: Kandasamy Saravanakumar, Myeong-Hyeon WangDocument9 pagesAdvanced Powder Technology: Kandasamy Saravanakumar, Myeong-Hyeon WangsilviaPas encore d'évaluation
- 3rd Summative Test in English - Q1Document4 pages3rd Summative Test in English - Q1Gessle GamirPas encore d'évaluation
- Acute Effects of Ayahuasca in A Juvenile Non-Human Primate Model of DepressionDocument9 pagesAcute Effects of Ayahuasca in A Juvenile Non-Human Primate Model of DepressionGonzaloBimontePas encore d'évaluation
- Roxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingD'EverandRoxane Gay & Everand Originals: My Year of Psychedelics: Lessons on Better LivingÉvaluation : 5 sur 5 étoiles5/5 (4)
- Lessons for Survival: Mothering Against “the Apocalypse”D'EverandLessons for Survival: Mothering Against “the Apocalypse”Évaluation : 5 sur 5 étoiles5/5 (2)
- The Comfort of Crows: A Backyard YearD'EverandThe Comfort of Crows: A Backyard YearÉvaluation : 4.5 sur 5 étoiles4.5/5 (23)
- The Secret Life of Lobsters: How Fishermen and Scientists Are Unraveling the Mysteries of Our Favorite CrustaceanD'EverandThe Secret Life of Lobsters: How Fishermen and Scientists Are Unraveling the Mysteries of Our Favorite CrustaceanPas encore d'évaluation
- The Storm of the Century: Tragedy, Heroism, Survival, and the Epic True Story of America's Deadliest Natural DisasterD'EverandThe Storm of the Century: Tragedy, Heroism, Survival, and the Epic True Story of America's Deadliest Natural DisasterPas encore d'évaluation