Vous aimerez peut-être aussi
- ParotidomégaliesDocument7 pagesParotidomégaliesOurouba HamriPas encore d'évaluation
- Dermatose ProfessionnelleDocument7 pagesDermatose ProfessionnellekgtdPas encore d'évaluation
- Etiologies Et Orientation Diagnostique Devant Un FlushDocument7 pagesEtiologies Et Orientation Diagnostique Devant Un FlushHenry TraoréPas encore d'évaluation
- Otite CanineDocument7 pagesOtite CanineAya DjouambiPas encore d'évaluation
- Candidas AlbicansDocument52 pagesCandidas AlbicansAbdou Latif RachidPas encore d'évaluation
- 8 1 - MastocytosesDocument17 pages8 1 - Mastocytoseselkabbadj mohammedPas encore d'évaluation
- Urticaires de Contact D'origine Professionnelle: en RésuméDocument12 pagesUrticaires de Contact D'origine Professionnelle: en Résuméfayssal bouakazPas encore d'évaluation
- Diagnostic en Parodontologie: I-IntroductionDocument10 pagesDiagnostic en Parodontologie: I-Introductionmohamed rayanPas encore d'évaluation
- Rfe Gestion Des Agents AntiplaquettairesDocument20 pagesRfe Gestion Des Agents Antiplaquettaireskarimsfar778Pas encore d'évaluation
- Dermato - Guide de Lexamen Clinique Et Du Diagnostic en DermatologieDocument294 pagesDermato - Guide de Lexamen Clinique Et Du Diagnostic en DermatologieNour CHPas encore d'évaluation
- Typage AmyloseDocument6 pagesTypage AmyloseHenry TraoréPas encore d'évaluation
- Pnds Sclerodermie WebDocument140 pagesPnds Sclerodermie WebParada AngelinePas encore d'évaluation
- Thèse D'exercice: Thèse Pour Le Diplôme D'etat de Docteur en PharmacieDocument95 pagesThèse D'exercice: Thèse Pour Le Diplôme D'etat de Docteur en PharmacieKhalil BelarbiPas encore d'évaluation
- Phéochromocytome Et ParagangliomeDocument9 pagesPhéochromocytome Et ParagangliomeAmina NibouchaPas encore d'évaluation
- 1-s2.0-S2667062323007134-mainDocument2 pages1-s2.0-S2667062323007134-mainTarik NajibPas encore d'évaluation
- P20113369 PDFDocument141 pagesP20113369 PDFYanni KhalfounePas encore d'évaluation
- TCC TrabalhoDocument24 pagesTCC TrabalhoCamile FerreiraPas encore d'évaluation
- Toxidermies: SciencedirectDocument15 pagesToxidermies: SciencedirectIbrahim AbdiPas encore d'évaluation
- Prise en Charge de La Dermatite AtopiqueDocument10 pagesPrise en Charge de La Dermatite AtopiquesofiaPas encore d'évaluation
- Toxoplasmose de La Femme EnceinteDocument12 pagesToxoplasmose de La Femme Enceintemaxime wotolPas encore d'évaluation
- Les Maladies de La Thyroïde (Jean-Louis Wémeau)Document184 pagesLes Maladies de La Thyroïde (Jean-Louis Wémeau)khriwa112Pas encore d'évaluation
- Ordonnances Mycologie MédicaleDocument16 pagesOrdonnances Mycologie Médicalehappy49100% (1)
- Avec La Participation De: (Manifestations Oculaires Exclues)Document27 pagesAvec La Participation De: (Manifestations Oculaires Exclues)Darryl LouisPas encore d'évaluation
- spf00003524Document42 pagesspf00003524ines.23dahouPas encore d'évaluation
- Phaeochromocytoma and Paraganglioma PDFDocument9 pagesPhaeochromocytoma and Paraganglioma PDFadmino blogoPas encore d'évaluation
- Dermato 2016 PDFDocument96 pagesDermato 2016 PDFjehane oussamaPas encore d'évaluation
- Scdpha T 2010 Boillot Amelie PDFDocument129 pagesScdpha T 2010 Boillot Amelie PDFSherley JeanPas encore d'évaluation
- FMC Fevrier 1Document8 pagesFMC Fevrier 1Ikramo CrPas encore d'évaluation
- Hello 2014Document11 pagesHello 2014khitasmoumenePas encore d'évaluation
- Physiopathologie de L'endométriose: A. Dumont, P. Collinet, B. Merlot, A. Abergel, C. Rubod, G. RobinDocument12 pagesPhysiopathologie de L'endométriose: A. Dumont, P. Collinet, B. Merlot, A. Abergel, C. Rubod, G. RobinMoncef Pecha100% (1)
- Couret 2019Document21 pagesCouret 2019Mohamed Wassim M'henniPas encore d'évaluation
- Cours de Parasitologie Et Mycologie CliniquesDocument204 pagesCours de Parasitologie Et Mycologie CliniquesMARCEL 'KENFACK100% (4)
- articlesJPA3 PDFDocument10 pagesarticlesJPA3 PDFBJ VeuxPas encore d'évaluation
- Les Maladies Sexuellement TransmissiblesDocument229 pagesLes Maladies Sexuellement TransmissiblesMedox007Pas encore d'évaluation
- MainDocument31 pagesMainSoumana BassirouPas encore d'évaluation
- Gougerot SjögrenDocument8 pagesGougerot Sjögrenalex mondomobePas encore d'évaluation
- 1 - Resumes Premiere Journee de L Internat Qualifiant en OdontostomatologieDocument20 pages1 - Resumes Premiere Journee de L Internat Qualifiant en OdontostomatologieTGPas encore d'évaluation
- CC Infect VZVDocument13 pagesCC Infect VZVmichifon yasminePas encore d'évaluation
- Le Diagnostic de La Maladie de Fabry Intérêt de L'enquête CliniqueDocument4 pagesLe Diagnostic de La Maladie de Fabry Intérêt de L'enquête Cliniquekhaledzoghbi16Pas encore d'évaluation
- 2005GRE17025 Freynet Laurent (1) (D) SODocument93 pages2005GRE17025 Freynet Laurent (1) (D) SOJonas SchreinemakersPas encore d'évaluation
- PNEDocument10 pagesPNERAKOTO NandrasanaPas encore d'évaluation
- Purpura Thrombopenique IdiopathiqueDocument69 pagesPurpura Thrombopenique IdiopathiqueAbdias ZOUNONPas encore d'évaluation
- Donadieu2017 PDFDocument11 pagesDonadieu2017 PDFMarouaBOUFLIJAPas encore d'évaluation
- Recommandations Françaises Pour La Prise en Charge de Pemphigus Jelti2019Document8 pagesRecommandations Françaises Pour La Prise en Charge de Pemphigus Jelti2019Faten HayderPas encore d'évaluation
- Strategies D'Enregistrement Des Produits de Sante: Une Interface Medicament-Dispositif MedicalDocument137 pagesStrategies D'Enregistrement Des Produits de Sante: Une Interface Medicament-Dispositif Medicalbouchra8blsPas encore d'évaluation
- Physiopathologie Et Traitement de La Maladie de StillDocument9 pagesPhysiopathologie Et Traitement de La Maladie de StillHenry TraoréPas encore d'évaluation
- RFE Etat de Mal Épileptique SRLF SFMUDocument46 pagesRFE Etat de Mal Épileptique SRLF SFMUAinane AyoubPas encore d'évaluation
- Dermatologie de La Diversité 2022Document284 pagesDermatologie de La Diversité 2022Mohamed takiy-eddine el moustafa SEGHIERPas encore d'évaluation
- Dermatologie Des Maladies D'organesDocument341 pagesDermatologie Des Maladies D'organesAhmed Dbb100% (4)
- 10 1016@j Admp 2011 07 006Document7 pages10 1016@j Admp 2011 07 006x-Abdelali Ait ouaichaPas encore d'évaluation
- abc-317172-45619-actualites_du_diagnostic_des_infections_parasitaires_et_fongiques-204216-u-4Document16 pagesabc-317172-45619-actualites_du_diagnostic_des_infections_parasitaires_et_fongiques-204216-u-4Sabi MaalemPas encore d'évaluation
- L'ophtalmologie ' A L'officine: B Er Enice CoquerelDocument213 pagesL'ophtalmologie ' A L'officine: B Er Enice CoquereldiamatkakaPas encore d'évaluation
- Actualites de La Dermatomyosite Juvenile PDFDocument6 pagesActualites de La Dermatomyosite Juvenile PDFRym MesfarPas encore d'évaluation
- J Annder 2019 10 027Document11 pagesJ Annder 2019 10 027Josué du237Pas encore d'évaluation
- Manifestaciones BucodentalesDocument5 pagesManifestaciones BucodentalessofysandovPas encore d'évaluation
- Proc To LogieDocument45 pagesProc To LogiealjallouliheshamPas encore d'évaluation
- EndoDocument14 pagesEndoZakariae Ibn AttyaPas encore d'évaluation
- 1997GRE17015 Hoarau Martine (0) SO Version DiffusionDocument155 pages1997GRE17015 Hoarau Martine (0) SO Version DiffusionAnou KalengaPas encore d'évaluation
- Interprétation Rapide De La Gazométrie Sanguine: La Gazométrie Sanguine En 4 ÉtapesD'EverandInterprétation Rapide De La Gazométrie Sanguine: La Gazométrie Sanguine En 4 ÉtapesPas encore d'évaluation
- TD N1 D'ANATOMIE ProfDocument3 pagesTD N1 D'ANATOMIE ProfAbdul Koudouss KonePas encore d'évaluation
- Les Voies de Conduction Au Niveau de La MoelleDocument38 pagesLes Voies de Conduction Au Niveau de La MoellesafemindPas encore d'évaluation
- Digestion Et Absorption PDFDocument6 pagesDigestion Et Absorption PDFrabab ouerdighiPas encore d'évaluation
- Samama 1964Document27 pagesSamama 1964Jimmy AndersPas encore d'évaluation
- 12 DMprocréationDocument9 pages12 DMprocréationDabé LaminePas encore d'évaluation
- Embryologie Humaine Polycopie PDF 1Document111 pagesEmbryologie Humaine Polycopie PDF 1LAFAYPas encore d'évaluation
- App UrinaireDocument8 pagesApp UrinaireHa NaàPas encore d'évaluation
- RappelDocument9 pagesRappeljaniaPas encore d'évaluation
- Développement Du Système ReproducteurDocument15 pagesDéveloppement Du Système ReproducteurGeorge CatalinPas encore d'évaluation
- TP2 PubertéDocument3 pagesTP2 PubertéAhane AssaniPas encore d'évaluation
- Lexique Vocabulaire Corps Humain en AnglaisDocument2 pagesLexique Vocabulaire Corps Humain en Anglaisquereur100% (1)
- Tissu Musculaire 1Document92 pagesTissu Musculaire 1Adonis Serghini100% (2)
- Anatomie Du Bloc Pancréas-Duodénum: Genu SuperiusDocument6 pagesAnatomie Du Bloc Pancréas-Duodénum: Genu SuperiusGhars AdamPas encore d'évaluation
- 2-Systeme MusculaireDocument6 pages2-Systeme MusculaireahlemmPas encore d'évaluation
- L'Appareil Digestif - 2Document24 pagesL'Appareil Digestif - 2El FaroukPas encore d'évaluation
- REIN Et HTADocument45 pagesREIN Et HTAdieynaba baPas encore d'évaluation
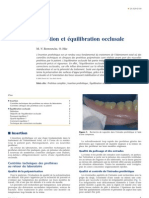
- Insertion Et ÉquilibrationDocument12 pagesInsertion Et Équilibrationgougou84Pas encore d'évaluation
- Segmentation Conduit Déférent - CopieDocument8 pagesSegmentation Conduit Déférent - CopieMARWA RAHOUHIPas encore d'évaluation
- Globules Blancs PR YAODocument63 pagesGlobules Blancs PR YAOJean Luc Konan100% (1)
- Margin Rajal-Ranap 2019Document235 pagesMargin Rajal-Ranap 2019rioPas encore d'évaluation
- HYPERTYROIDIE Cours 2023 FacDocument7 pagesHYPERTYROIDIE Cours 2023 Facchristnelle chiruda mafoutaPas encore d'évaluation
- Transfusion SanguineDocument159 pagesTransfusion SanguineIsmael Dk100% (1)
- Shémas Hemostase 06Document10 pagesShémas Hemostase 06coachboubouPas encore d'évaluation
- 09 La Communication Nerveuse PDFDocument11 pages09 La Communication Nerveuse PDFwejdenPas encore d'évaluation
- L Appreil Reproducteur de L HommeDocument88 pagesL Appreil Reproducteur de L HommeFouzia ChouliPas encore d'évaluation
- Rapport Bioanalyses MedicalesDocument26 pagesRapport Bioanalyses MedicalesBrahim Omari100% (1)
- Corrige 472459Document80 pagesCorrige 472459PFEPas encore d'évaluation
- Texte À Trous. L'appareil DigestifDocument1 pageTexte À Trous. L'appareil DigestifIrene PG100% (1)
- Néphrectomie Simple Et Néphrectomie Elargie Pour CancerDocument28 pagesNéphrectomie Simple Et Néphrectomie Elargie Pour Cancerdjasmi mohamedPas encore d'évaluation
- Les Methodes en MusculationDocument8 pagesLes Methodes en MusculationerrajiePas encore d'évaluation