Vous aimerez peut-être aussi
- Type 1 Diabetes A Chronic Anti-Self-Inflammatory ResponseDocument10 pagesType 1 Diabetes A Chronic Anti-Self-Inflammatory ResponseIda YulianaPas encore d'évaluation
- Inmunobiologia CA RenalDocument7 pagesInmunobiologia CA RenalDelia Lucia Escola GomezPas encore d'évaluation
- Tumor Necrosis Factor and Anti-Tumor Necrosis Factor TherapiesDocument13 pagesTumor Necrosis Factor and Anti-Tumor Necrosis Factor TherapiesAgus PrimaPas encore d'évaluation
- CD4 Tregs and Immune Control: Review SeriesDocument9 pagesCD4 Tregs and Immune Control: Review SeriesFelipe Gálvez JirónPas encore d'évaluation
- Anti-TNF Therapy in Crohn's Disease: Molecular SciencesDocument21 pagesAnti-TNF Therapy in Crohn's Disease: Molecular SciencesLuiz Filipe BezerraPas encore d'évaluation
- MHC and Disease Susceptibility (Type 1 Diabetes As Example)Document8 pagesMHC and Disease Susceptibility (Type 1 Diabetes As Example)FATbovenPas encore d'évaluation
- Epigenetics & Modulation of ImmunityDocument6 pagesEpigenetics & Modulation of ImmunityAnonymous pFdFBlXO00Pas encore d'évaluation
- Nri 1936 ArtigoDocument13 pagesNri 1936 ArtigoGlauce L TrevisanPas encore d'évaluation
- T Cells in Health and DiseaseDocument50 pagesT Cells in Health and Diseaseygilad9139Pas encore d'évaluation
- Fimmu 12 681062Document13 pagesFimmu 12 681062Omnia AshrafPas encore d'évaluation
- Targeting T Cell Activation in Immuno-Oncology: S.D. Saibil and P.S. OhashiDocument8 pagesTargeting T Cell Activation in Immuno-Oncology: S.D. Saibil and P.S. OhashiNBME testPas encore d'évaluation
- Cytokines Immunotherapy-Bayry 2007 NTR RVW ImmnlDocument2 pagesCytokines Immunotherapy-Bayry 2007 NTR RVW Immnl2shankarPas encore d'évaluation
- Activating Immunity To Fight A FoeDocument3 pagesActivating Immunity To Fight A FoeGabriela ArvelaezPas encore d'évaluation
- 10 1016@j TCB 2020 06 003Document10 pages10 1016@j TCB 2020 06 003Maria Aponte RuizPas encore d'évaluation
- Cellular and Molecular Pathophysiology of Type 1 Diabetes MellitusDocument8 pagesCellular and Molecular Pathophysiology of Type 1 Diabetes Mellitussofia.rvvvPas encore d'évaluation
- Immunometabolism of T Cells and NK Cells: Metabolic Control of Effector and Regulatory FunctionDocument16 pagesImmunometabolism of T Cells and NK Cells: Metabolic Control of Effector and Regulatory FunctionDHRVAT15Pas encore d'évaluation
- Nita - Ming O Li A Master of All T Cell TradesDocument24 pagesNita - Ming O Li A Master of All T Cell TradesYuwan RamjadaPas encore d'évaluation
- Fimmu 13 816005Document14 pagesFimmu 13 816005emmanuel AndemPas encore d'évaluation
- Points of Control in Inflammation: InsightDocument7 pagesPoints of Control in Inflammation: InsightAsish GeiorgePas encore d'évaluation
- CD39 A Promising Target in Oncology PDFDocument9 pagesCD39 A Promising Target in Oncology PDFGeorge Sebastian AntonyPas encore d'évaluation
- Roychoudhuri 2015Document11 pagesRoychoudhuri 2015MaríaFernandaEcheverryPas encore d'évaluation
- Co-signal Molecules in T Cell Activation: Immune Regulation in Health and DiseaseD'EverandCo-signal Molecules in T Cell Activation: Immune Regulation in Health and DiseaseMiyuki AzumaPas encore d'évaluation
- 17-Cancer and The Immune SystemDocument25 pages17-Cancer and The Immune SystemRami AlhilliPas encore d'évaluation
- 2019 - Regulatory T Cells Peptide-MHC Class II From Dendritic CellsDocument19 pages2019 - Regulatory T Cells Peptide-MHC Class II From Dendritic Cells1262615286Pas encore d'évaluation
- Ced 13795Document7 pagesCed 13795Mega MemoryPas encore d'évaluation
- Fimmu 11 622810Document9 pagesFimmu 11 622810Rovin RamphalPas encore d'évaluation
- Cytokine Pathways and Joint Inflammation in Rheumatoid ArthritisDocument10 pagesCytokine Pathways and Joint Inflammation in Rheumatoid ArthritisNICOLASPas encore d'évaluation
- T Cell ApoptosisDocument9 pagesT Cell ApoptosisVidho RiveraPas encore d'évaluation
- Struma Nodosa 2Document10 pagesStruma Nodosa 2dewi rositaPas encore d'évaluation
- Anti InflammatoryDocument11 pagesAnti InflammatoryMazlan SijifPas encore d'évaluation
- Dejaco-Et-Al 06 - (Imbalance of Regulatory T Cells in Human Autoimmune Diseases)Document12 pagesDejaco-Et-Al 06 - (Imbalance of Regulatory T Cells in Human Autoimmune Diseases)Luana DiEmmePas encore d'évaluation
- Anti-IL-21 Monoclonal Antibody Combined With Liraglutide Effectively Reverses Established Hyperglycemia in Mouse Models of Type 1 Diabetes.Document10 pagesAnti-IL-21 Monoclonal Antibody Combined With Liraglutide Effectively Reverses Established Hyperglycemia in Mouse Models of Type 1 Diabetes.rafaelpires85Pas encore d'évaluation
- 100 Years of InsuinmDocument11 pages100 Years of InsuinmJoão Guilherme HazinPas encore d'évaluation
- 2018 - STAT3 in Regulatory T Cells Are PD-1 Blockade in Resected Metastatic MelanomaDocument12 pages2018 - STAT3 in Regulatory T Cells Are PD-1 Blockade in Resected Metastatic Melanoma1262615286Pas encore d'évaluation
- 2001 - Regulation of IFN-γ signaling is essential for the cytotoxic activity of CD8+Document10 pages2001 - Regulation of IFN-γ signaling is essential for the cytotoxic activity of CD8+顏世隆Pas encore d'évaluation
- 2 - Celulas T Gama DeltaDocument8 pages2 - Celulas T Gama DeltaDan RyuPas encore d'évaluation
- 1a PDFDocument10 pages1a PDFAndrea Il MiodoPas encore d'évaluation
- Jenis Situokin Dan Relevansi Klinis. Ganong's Review of Medical Physiology, 25 Ed. 2016Document1 pageJenis Situokin Dan Relevansi Klinis. Ganong's Review of Medical Physiology, 25 Ed. 2016Iradatullah SuyutiPas encore d'évaluation
- E003293 FullDocument16 pagesE003293 FullNeni Rahawarin03Pas encore d'évaluation
- 44 pdl1Document3 pages44 pdl1bawoji1763Pas encore d'évaluation
- Wang 2003Document24 pagesWang 2003Xena LavedaPas encore d'évaluation
- Inmunología 4Document8 pagesInmunología 4Valentina PereiraPas encore d'évaluation
- Science - Aaa4971 2Document7 pagesScience - Aaa4971 2Alejandro Zuñiga NaterasPas encore d'évaluation
- Mesenchymal Stem Cells in COVID-19 - A Journey From Bench To BedsideDocument12 pagesMesenchymal Stem Cells in COVID-19 - A Journey From Bench To Bedsidediana.alyPas encore d'évaluation
- Gmbar ImunDocument10 pagesGmbar ImunVera FitrianaPas encore d'évaluation
- Fonc 08 00086Document14 pagesFonc 08 00086Siska HarapanPas encore d'évaluation
- SGD 23 - Immunology 3Document4 pagesSGD 23 - Immunology 3John RayPas encore d'évaluation
- 1 s2.0 S2405579421000036 MainDocument8 pages1 s2.0 S2405579421000036 MainNurul HusnaPas encore d'évaluation
- Tubulointerstitial Renal Disease - Curr Opin Nephrol HypertenDocument11 pagesTubulointerstitial Renal Disease - Curr Opin Nephrol HypertenErnestoPas encore d'évaluation
- 10 1016@j Immuni 2020 10 002Document20 pages10 1016@j Immuni 2020 10 002Licia TorresPas encore d'évaluation
- DR Zuhair Sabbagh - Biologics in Rheumatoid Arthritis - Medics Index MemberDocument7 pagesDR Zuhair Sabbagh - Biologics in Rheumatoid Arthritis - Medics Index MemberMedicsindex Telepin Slidecase100% (2)
- Artigo ImunologiaDocument13 pagesArtigo ImunologiapaulamonteiromedPas encore d'évaluation
- T-Cell Exhaustion and Barriers to ImmunotherapyDocument24 pagesT-Cell Exhaustion and Barriers to Immunotherapyro111111Pas encore d'évaluation
- CYTOKINESDocument31 pagesCYTOKINESSaifi Alam100% (4)
- Tumor necrosis factor and cancer, buddies or foes?Document14 pagesTumor necrosis factor and cancer, buddies or foes?hoangphuong08101992Pas encore d'évaluation
- INIM0011A6UC - 2021 - B1 Paper HandoutDocument11 pagesINIM0011A6UC - 2021 - B1 Paper HandoutASDPas encore d'évaluation
- Reviews: Pancreatic β-cells in type 1 and type 2 diabetes mellitus: different pathways to failureDocument14 pagesReviews: Pancreatic β-cells in type 1 and type 2 diabetes mellitus: different pathways to failureJESSICA CELESTE LUNA PEÑAPas encore d'évaluation
- TNFR, TRAF2, NF-κB mRNA Levels of Glioblastoma Multiforme Cells Treated by Conditioned Medium of Umbilical Cord-derived Mesenchymal Stem CellsDocument8 pagesTNFR, TRAF2, NF-κB mRNA Levels of Glioblastoma Multiforme Cells Treated by Conditioned Medium of Umbilical Cord-derived Mesenchymal Stem CellsVinnie Juliana YonatanPas encore d'évaluation
- Tumor Immune Microenvironment in Cancer Progression and Cancer TherapyD'EverandTumor Immune Microenvironment in Cancer Progression and Cancer TherapyPawel KalinskiPas encore d'évaluation
- Fast Facts: Managing immune-related Adverse Events in Oncology: Early recognition, prompt intervention, effective managementD'EverandFast Facts: Managing immune-related Adverse Events in Oncology: Early recognition, prompt intervention, effective managementPas encore d'évaluation
- MCAT 2015 OutlineDocument128 pagesMCAT 2015 OutlinequirkyheadbandsPas encore d'évaluation
- 2018 REVIEW: of Physician and Advanced Practitioner Recruiting IncentivesDocument56 pages2018 REVIEW: of Physician and Advanced Practitioner Recruiting IncentivesBen BorokhovskyPas encore d'évaluation
- MCAT Essentials 2016 - FinalDocument42 pagesMCAT Essentials 2016 - FinalBenjamin PhuaPas encore d'évaluation
- Results and Data SMS 2019Document160 pagesResults and Data SMS 2019Ben BorokhovskyPas encore d'évaluation
- CT HistologyDocument1 pageCT HistologyBen BorokhovskyPas encore d'évaluation
- (2001) Reversal of Established Autoimmune Diabetes by Restoration of Endogenous β Cell FunctionDocument10 pages(2001) Reversal of Established Autoimmune Diabetes by Restoration of Endogenous β Cell FunctionBen BorokhovskyPas encore d'évaluation
- CH 14Document11 pagesCH 14Ben BorokhovskyPas encore d'évaluation
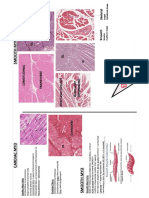
- Smooth Cardiac Histology SlidesDocument1 pageSmooth Cardiac Histology SlidesBen BorokhovskyPas encore d'évaluation
- (2001) Reversal of Established Autoimmune Diabetes by Restoration of Endogenous β Cell FunctionDocument10 pages(2001) Reversal of Established Autoimmune Diabetes by Restoration of Endogenous β Cell FunctionBen BorokhovskyPas encore d'évaluation
- Skeletal Myo HistologyDocument1 pageSkeletal Myo HistologyBen BorokhovskyPas encore d'évaluation
- Battleplan REALM QUEST PDFDocument2 pagesBattleplan REALM QUEST PDFBen BorokhovskyPas encore d'évaluation
- Hardy-Ramanujan Asymptotic Partition Formula approximates number of integer partitionsDocument1 pageHardy-Ramanujan Asymptotic Partition Formula approximates number of integer partitionsBen BorokhovskyPas encore d'évaluation
- (2015) Low Levels of C-Peptide Have Clinical Significance For Established Type 1 DiabetesDocument8 pages(2015) Low Levels of C-Peptide Have Clinical Significance For Established Type 1 DiabetesBen BorokhovskyPas encore d'évaluation
- (2003) Islet Regeneration During The Reversal of Autoimmune Diabetes in NOD MiceDocument6 pages(2003) Islet Regeneration During The Reversal of Autoimmune Diabetes in NOD MiceBen BorokhovskyPas encore d'évaluation
- Hardy-Ramanujan Asymptotic Partition Formula approximates number of integer partitionsDocument1 pageHardy-Ramanujan Asymptotic Partition Formula approximates number of integer partitionsBen BorokhovskyPas encore d'évaluation
- Charting Outcomes in The Match 2018 Seniors PDFDocument220 pagesCharting Outcomes in The Match 2018 Seniors PDFgus_lionsPas encore d'évaluation
- SEX - Decoded and Explained - Free AdvancedDocument194 pagesSEX - Decoded and Explained - Free AdvancedBen Borokhovsky100% (1)
- (2010) TNF Receptor 2 Pathway - Drug Target For Autoimmune DiseasesDocument12 pages(2010) TNF Receptor 2 Pathway - Drug Target For Autoimmune DiseasesBen BorokhovskyPas encore d'évaluation
- (2015) Low Levels of C-Peptide Have Clinical Significance For Established Type 1 DiabetesDocument8 pages(2015) Low Levels of C-Peptide Have Clinical Significance For Established Type 1 DiabetesBen BorokhovskyPas encore d'évaluation
- (2007) The Promise of Hox11 + Stem Cells of The Spleen For Treating Autoimmune DiseasesDocument10 pages(2007) The Promise of Hox11 + Stem Cells of The Spleen For Treating Autoimmune DiseasesBen BorokhovskyPas encore d'évaluation
- JournalPone0041756 (1) Clinical Trials Phase 1Document16 pagesJournalPone0041756 (1) Clinical Trials Phase 1Anuj MittalPas encore d'évaluation
- (2001) Reversal of Established Autoimmune Diabetes by Restoration of Endogenous β Cell FunctionDocument10 pages(2001) Reversal of Established Autoimmune Diabetes by Restoration of Endogenous β Cell FunctionBen BorokhovskyPas encore d'évaluation
- (2003) Islet Regeneration During The Reversal of Autoimmune Diabetes in NOD MiceDocument6 pages(2003) Islet Regeneration During The Reversal of Autoimmune Diabetes in NOD MiceBen BorokhovskyPas encore d'évaluation
- (2010) TNF Receptor 2 Pathway - Drug Target For Autoimmune DiseasesDocument12 pages(2010) TNF Receptor 2 Pathway - Drug Target For Autoimmune DiseasesBen BorokhovskyPas encore d'évaluation
- (2015) Low Levels of C-Peptide Have Clinical Significance For Established Type 1 DiabetesDocument8 pages(2015) Low Levels of C-Peptide Have Clinical Significance For Established Type 1 DiabetesBen BorokhovskyPas encore d'évaluation
- (2001) Reversal of Established Autoimmune Diabetes by Restoration of Endogenous β Cell FunctionDocument10 pages(2001) Reversal of Established Autoimmune Diabetes by Restoration of Endogenous β Cell FunctionBen BorokhovskyPas encore d'évaluation
- 101 Sext Phrases GuideDocument7 pages101 Sext Phrases GuideBen Borokhovsky100% (1)
- QLAeBook-2nd Edition PDFDocument34 pagesQLAeBook-2nd Edition PDFBen BorokhovskyPas encore d'évaluation
- اهم اسئلة 2Document32 pagesاهم اسئلة 2Adam JerusalemPas encore d'évaluation
- Barsanalina - Assignment For Chapter 16 Adaptive ImmunityDocument4 pagesBarsanalina - Assignment For Chapter 16 Adaptive ImmunityRalph B. BarsanalinaPas encore d'évaluation
- Borrador Último Jenny Laia Aste Teresa 200723Document19 pagesBorrador Último Jenny Laia Aste Teresa 200723Teresa RibaltaPas encore d'évaluation
- Fimmu 09 00328Document11 pagesFimmu 09 00328miss betawiPas encore d'évaluation
- Chapter 6 - Immune Diseases (Part I) (Robbins and Cotran Pathologic Basis of Disease)Document6 pagesChapter 6 - Immune Diseases (Part I) (Robbins and Cotran Pathologic Basis of Disease)Ernie G. Bautista II, RN, MD90% (21)
- PG M.sc. Zoology 35022 ImmunologyDocument241 pagesPG M.sc. Zoology 35022 ImmunologyJo FighizPas encore d'évaluation
- Understanding AIDS: A Guide to its Causes, Symptoms, Diagnosis and PreventionDocument27 pagesUnderstanding AIDS: A Guide to its Causes, Symptoms, Diagnosis and PreventionNikhil BijuPas encore d'évaluation
- Haematology: Lecture NotesDocument4 pagesHaematology: Lecture NotesjonasPas encore d'évaluation
- MHC diversity and antigen presentation pathwaysDocument2 pagesMHC diversity and antigen presentation pathwaysyolo51Pas encore d'évaluation
- ImmunologyDocument91 pagesImmunologyNicole FigueroaPas encore d'évaluation
- Systemic Lupus Erythematosus 2012Document564 pagesSystemic Lupus Erythematosus 2012Dan MunteanuPas encore d'évaluation
- Immunity Question Paper For A Level BiologyDocument2 pagesImmunity Question Paper For A Level BiologyJansi Angel100% (1)
- Tumor Immunology: Understanding Immune Reactivity and ApproachesDocument20 pagesTumor Immunology: Understanding Immune Reactivity and ApproachesMarl EstradaPas encore d'évaluation
- Immunoinformatics Notes2019Document20 pagesImmunoinformatics Notes2019Anamitra SenPas encore d'évaluation
- Test Bank For Basic Immunology 3rd Edition Abul K Abbas Full DownloadDocument6 pagesTest Bank For Basic Immunology 3rd Edition Abul K Abbas Full Downloadbrianbell06021999pfa100% (34)
- Overview of The Immune System 2020Document31 pagesOverview of The Immune System 2020mehakPas encore d'évaluation
- Chapter 17: Biopsychology of Emotion, Stress, and Health Stress and HealthDocument3 pagesChapter 17: Biopsychology of Emotion, Stress, and Health Stress and HealthRashia LubuguinPas encore d'évaluation
- Chapter 6 - Diseases of The Immune SystemDocument12 pagesChapter 6 - Diseases of The Immune SystemAgnieszka WisniewskaPas encore d'évaluation
- Alopecia Totalis Following HFMDDocument3 pagesAlopecia Totalis Following HFMDVilt VilPas encore d'évaluation
- David Gate CV 9-19-18Document5 pagesDavid Gate CV 9-19-18David GatePas encore d'évaluation
- CL1 HaemopoiesisDocument41 pagesCL1 HaemopoiesisSamuel George100% (1)
- Biology - Assignment 5 - Pathogens and The Immune System - ReportDocument10 pagesBiology - Assignment 5 - Pathogens and The Immune System - Reportapi-585453254Pas encore d'évaluation
- 3 s2.0 B9780128185612000047 MainDocument19 pages3 s2.0 B9780128185612000047 Maindimas rifqiPas encore d'évaluation
- Epstein-Barr Virus, The Immune System, and Associated DiseasesDocument5 pagesEpstein-Barr Virus, The Immune System, and Associated DiseasesArlen JamesPas encore d'évaluation
- CRISPR screens decode cancer cell pathways that trigger γδ T cell detectionDocument31 pagesCRISPR screens decode cancer cell pathways that trigger γδ T cell detectionygilad9139Pas encore d'évaluation
- MitochondriaDocument6 pagesMitochondriakevin smithPas encore d'évaluation
- Anatomy & Physiology: Essentials ofDocument69 pagesAnatomy & Physiology: Essentials ofclyde i am100% (1)
- IMTX PatentDocument72 pagesIMTX PatentCharles GrossPas encore d'évaluation
- Renal Immunology PDFDocument108 pagesRenal Immunology PDFMayra Elizabeth RodriguezPas encore d'évaluation
- MR Supple All 2014Document429 pagesMR Supple All 2014Regita Ayu LestariPas encore d'évaluation