Vous aimerez peut-être aussi
- Oficio de Reporte de CuentaDocument1 pageOficio de Reporte de CuentaRafo CastroPas encore d'évaluation
- Modelo de Escrito de AUTORIZO PARA REVISION DE EXPEDIENTESDocument1 pageModelo de Escrito de AUTORIZO PARA REVISION DE EXPEDIENTESRafo CastroPas encore d'évaluation
- Recurso de Apelación Contra PapeletaDocument13 pagesRecurso de Apelación Contra PapeletaRafo Castro100% (2)
- Informe de Sistematica n5Document14 pagesInforme de Sistematica n5Rafo CastroPas encore d'évaluation
- Libro de BiologiaDocument243 pagesLibro de BiologiaRafo CastroPas encore d'évaluation
- Clase Mixosporidea y MicrosporideaDocument8 pagesClase Mixosporidea y MicrosporideaRafo CastroPas encore d'évaluation
- Jurisprudencia LaboralDocument1 pageJurisprudencia LaboralRafo CastroPas encore d'évaluation
- ALGODONDocument12 pagesALGODONRafo CastroPas encore d'évaluation
- Memoria Topografia S.F.asisDocument13 pagesMemoria Topografia S.F.asisRoberto Jhony Huaripata RomeroPas encore d'évaluation
- El Desarrollo de La Hipótesis de La Deriva Continental Se Le Atribuye A Alfred WegenerDocument2 pagesEl Desarrollo de La Hipótesis de La Deriva Continental Se Le Atribuye A Alfred WegenerJohana Talavera67% (9)
- Reseña Histórica de BiscucuyDocument4 pagesReseña Histórica de BiscucuyMaria Barazarte100% (2)
- Est. Hidrologico - Canal SaavedraDocument37 pagesEst. Hidrologico - Canal SaavedracesarPas encore d'évaluation
- Trampas Psicologicas en La Toma de DecisionesDocument6 pagesTrampas Psicologicas en La Toma de Decisioneslauramedina17Pas encore d'évaluation
- Introduccion A La Propagacion de Plantas (Mani)Document18 pagesIntroduccion A La Propagacion de Plantas (Mani)Jill Engell Vasquez Calero0% (1)
- AristótelesDocument6 pagesAristótelesחוארז עומרPas encore d'évaluation
- Proyecto de Investigacion Incidencia Del Factor de Riesgo Biologico - Prof AnaDocument49 pagesProyecto de Investigacion Incidencia Del Factor de Riesgo Biologico - Prof AnaLUIS ERNESTO REINA ROMERO100% (2)
- Problema 2Document1 pageProblema 2Jorge Vesga SuarezPas encore d'évaluation
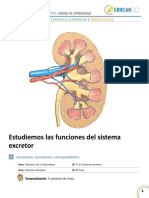
- RiñonDocument12 pagesRiñonMaria Luisa Correa GalvezPas encore d'évaluation
- Corteza ContinentalDocument6 pagesCorteza ContinentalJuanPas encore d'évaluation
- El Efecto Dunning-KrugerDocument4 pagesEl Efecto Dunning-Krugerjibanezo3216100% (2)
- Redes TroficasDocument17 pagesRedes TroficasJonathan Paul Otoya BarturenPas encore d'évaluation
- ColtánDocument23 pagesColtánFco Javier Benítez RubioPas encore d'évaluation
- Glaciar TicllaDocument40 pagesGlaciar TicllaDaniel LoPas encore d'évaluation
- DiarioLuna CarlotaRamosAbreu2BACHA.Document82 pagesDiarioLuna CarlotaRamosAbreu2BACHA.Carlota RamosPas encore d'évaluation
- Diseno de Zapatas AisladasDocument14 pagesDiseno de Zapatas AisladasMiriam Gimena Choque AruquipaPas encore d'évaluation
- Actividad HexoquinasaDocument12 pagesActividad HexoquinasaJuan Domingo Davila GilPas encore d'évaluation
- Cementacion de Cobre-CrisDocument6 pagesCementacion de Cobre-CrisALEXANDER NAZARIO ACEVEDO SINCHEPas encore d'évaluation
- Historia de La MineralogíaDocument2 pagesHistoria de La MineralogíaSergio Hernandez AguilarPas encore d'évaluation
- Coordenadas PolaresDocument2 pagesCoordenadas PolaresAlexys RuizPas encore d'évaluation
- Guia de Construccion - BaharequeDocument28 pagesGuia de Construccion - BaharequeMajo GrajedaPas encore d'évaluation
- Guia Unitips PDFDocument36 pagesGuia Unitips PDFNicole Yañez67% (6)
- Guia #12. SocialesDocument4 pagesGuia #12. SocialesRouss Marie100% (1)
- LanaDocument10 pagesLanaRober OrtizPas encore d'évaluation
- Silabo Estomatologia Preventiva 2007Document5 pagesSilabo Estomatologia Preventiva 2007Luis sanchezPas encore d'évaluation
- Physio ExDocument15 pagesPhysio Exissa2897Pas encore d'évaluation
- Determinacion Del Carbono Orgcanico en SedimentosDocument4 pagesDeterminacion Del Carbono Orgcanico en SedimentosDavid Sergio Valdés LozanoPas encore d'évaluation
- Ficha Tecnica YogurtDocument1 pageFicha Tecnica YogurtPepe VasquezPas encore d'évaluation
- Comprensión de Lectura SextoDocument11 pagesComprensión de Lectura SextoCarolinaMarimanPas encore d'évaluation