Vous aimerez peut-être aussi
- Alimentacion SaludablepdfesteticaDocument9 pagesAlimentacion SaludablepdfesteticaNadia ReineroPas encore d'évaluation
- CRIOLIPOLISISTEORIADocument6 pagesCRIOLIPOLISISTEORIANadia ReineroPas encore d'évaluation
- Reglamento de Racata 1,2Document2 pagesReglamento de Racata 1,2Nadia ReineroPas encore d'évaluation
- Criolipolisis CursoDocument5 pagesCriolipolisis CursoNadia ReineroPas encore d'évaluation
- Madero MasterDocument44 pagesMadero MasterNadia ReineroPas encore d'évaluation
- Sugerencias de Uso Crio Portable 2019 NizaDocument4 pagesSugerencias de Uso Crio Portable 2019 NizaNadia ReineroPas encore d'évaluation
- Alumno QuinterosDocument1 pageAlumno QuinterosNadia ReineroPas encore d'évaluation
- Vademecum Lista CosmeticaDocument23 pagesVademecum Lista CosmeticaNadia Reinero100% (1)
- Toma de MedidasDocument3 pagesToma de MedidasNadia ReineroPas encore d'évaluation
- Molde Base Espalda. (J)Document6 pagesMolde Base Espalda. (J)Nadia ReineroPas encore d'évaluation
- Peelings Precios 01nov2022Document2 pagesPeelings Precios 01nov2022Nadia ReineroPas encore d'évaluation
- Tecnica Del Masaje Terapeutico Gua ShaDocument3 pagesTecnica Del Masaje Terapeutico Gua ShaNadia ReineroPas encore d'évaluation
- Libro de Ejercicios Akasicos Nivel 1 PDFDocument393 pagesLibro de Ejercicios Akasicos Nivel 1 PDFMarta Moena96% (25)
- Prove Edo ResDocument2 pagesProve Edo ResNadia ReineroPas encore d'évaluation
- Master en CosmiatriaDocument1 pageMaster en CosmiatriaNadia ReineroPas encore d'évaluation
- Partida 279708Document2 pagesPartida 279708Nadia ReineroPas encore d'évaluation
- Presentacion BB GlowDocument20 pagesPresentacion BB GlowNadia ReineroPas encore d'évaluation
- TRABAJO PRACTICO NUMERO 1 Modulo 1Document1 pageTRABAJO PRACTICO NUMERO 1 Modulo 1Nadia ReineroPas encore d'évaluation
- BODYTEORIA1Document9 pagesBODYTEORIA1Nadia ReineroPas encore d'évaluation
- PdfteoriaelectroesteticaDocument17 pagesPdfteoriaelectroesteticaNadia ReineroPas encore d'évaluation
- Pedido 1Document1 pagePedido 1Nadia ReineroPas encore d'évaluation
- EspatulapdfDocument6 pagesEspatulapdfNadia ReineroPas encore d'évaluation
- Master Teoria 1Document36 pagesMaster Teoria 1Nadia ReineroPas encore d'évaluation
- Microdermoabrasion ElectroporadorDocument4 pagesMicrodermoabrasion ElectroporadorNadia ReineroPas encore d'évaluation
- TAPAPEELINGDocument1 pageTAPAPEELINGNadia ReineroPas encore d'évaluation
- Progama Masoterapia 2022Document1 pageProgama Masoterapia 2022Nadia ReineroPas encore d'évaluation
- FotospeelingDocument13 pagesFotospeelingNadia ReineroPas encore d'évaluation
- AtelierDocument6 pagesAtelierNadia ReineroPas encore d'évaluation
- HilosteoriaDocument8 pagesHilosteoriaNadia ReineroPas encore d'évaluation
- COSME2022PROGRAMADocument1 pageCOSME2022PROGRAMANadia ReineroPas encore d'évaluation
- Cuadro Comparativo Entre PsicoterapiasDocument3 pagesCuadro Comparativo Entre PsicoterapiasHellen RfPas encore d'évaluation
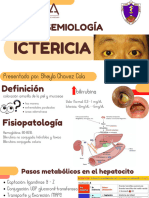
- ICTERICIADocument12 pagesICTERICIASHEYLA FIORELA CHAVEZ CALAPas encore d'évaluation
- Laboratorio de Edafologia 9 SalinidadDocument4 pagesLaboratorio de Edafologia 9 SalinidadJeferDavila0% (1)
- Estilos de Aprendizaje InfografiaDocument1 pageEstilos de Aprendizaje InfografiaPaula VillegasPas encore d'évaluation
- Trabajo Practico No 3. Alternativas de Aprovechamiento y ValorizacionDocument3 pagesTrabajo Practico No 3. Alternativas de Aprovechamiento y ValorizacionAndrea LadinoPas encore d'évaluation
- Unidad - 2 - 1 Diplomado PDFDocument67 pagesUnidad - 2 - 1 Diplomado PDFCarolina Castillo HidalgoPas encore d'évaluation
- Fusibles y Relés para Audi A4 - S4 (B8 - 8K 2008-2016) - Diagramas de Caja de FusiblesDocument16 pagesFusibles y Relés para Audi A4 - S4 (B8 - 8K 2008-2016) - Diagramas de Caja de FusiblesRuben David Ibañez RetamalPas encore d'évaluation
- Tema 3 Registros en La História ClínicaDocument8 pagesTema 3 Registros en La História ClínicaKellycita Barreto AlavaPas encore d'évaluation
- Carta The FitzgeraldDocument4 pagesCarta The FitzgeraldAwpov XizpaPas encore d'évaluation
- Ensayo Sobre La Aplicacion de Quimica en La IndustriaDocument11 pagesEnsayo Sobre La Aplicacion de Quimica en La Industriajhudith torresPas encore d'évaluation
- Contextualizaciòn Del CasoDocument2 pagesContextualizaciòn Del CasoAngélica LealPas encore d'évaluation
- Formato de Contenido de Plan de Gestión Integral de Desechos SanitariosDocument4 pagesFormato de Contenido de Plan de Gestión Integral de Desechos SanitariosPaoPilcoVirePas encore d'évaluation
- Miedo Al ExitoDocument15 pagesMiedo Al ExitoTONYPas encore d'évaluation
- Bloque Quimica 3ro BGUDocument31 pagesBloque Quimica 3ro BGUCarlos Oswaldo García ParedesPas encore d'évaluation
- Últimas Investigaciones Sobre El TDAH (Oct 09)Document2 pagesÚltimas Investigaciones Sobre El TDAH (Oct 09)Beatriz Peroy VielaPas encore d'évaluation
- Informe 3 Determinacion Del Diametro Molecular de RamosDocument19 pagesInforme 3 Determinacion Del Diametro Molecular de RamosRAMOS OLLACHICA MARYORI JENNIFERPas encore d'évaluation
- EXTRACCIÓNDocument5 pagesEXTRACCIÓNGeorges FloresPas encore d'évaluation
- 1 TDR Camara de Empalme y MedicionDocument23 pages1 TDR Camara de Empalme y MedicionAnthony Soto MegoPas encore d'évaluation
- Esquema de Manejo Ambiental TutaDocument29 pagesEsquema de Manejo Ambiental TutaLuis Fernando PerdomoPas encore d'évaluation
- Elaboración de Juguetes de Material Reciclado para Los Niños Del Servicio de PediatríaDocument35 pagesElaboración de Juguetes de Material Reciclado para Los Niños Del Servicio de PediatríaDeborah Valdez BellidoPas encore d'évaluation
- Mapas 26Document5 pagesMapas 26FPMPas encore d'évaluation
- Darapel 1 PDFDocument2 pagesDarapel 1 PDFkarina ortega0% (1)
- Evaluación Del AguaDocument4 pagesEvaluación Del AguaBarbara JaraPas encore d'évaluation
- Mapa DemandaDocument1 pageMapa DemandaLuis Enrique Vera CamposPas encore d'évaluation
- Informe de Transito Estacion El AeropuertoDocument5 pagesInforme de Transito Estacion El AeropuertoMEGATETOMPas encore d'évaluation
- Quimica 4 ElementosDocument7 pagesQuimica 4 ElementosArnold RamirezPas encore d'évaluation
- Servidumbres de AcueductoDocument7 pagesServidumbres de AcueductoEdgardo GaleanoPas encore d'évaluation
- Inf. Viscosidad y Tensión Superficial TexacaDocument14 pagesInf. Viscosidad y Tensión Superficial TexacaDenilson ChaconPas encore d'évaluation
- HidroestàticaDocument12 pagesHidroestàticaTomi AlbouyPas encore d'évaluation
- Actualización Covid-19 en GV, Vie 7 de Enero, 2022 GV Update Covid-19 - Fri Jan 07, 2022Document4 pagesActualización Covid-19 en GV, Vie 7 de Enero, 2022 GV Update Covid-19 - Fri Jan 07, 2022Carlos MirandaPas encore d'évaluation