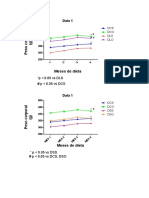
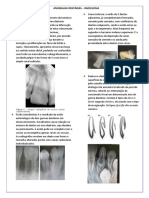
Vous aimerez peut-être aussi
- Ensino sinais pontuação 7a classeDocument36 pagesEnsino sinais pontuação 7a classecaetano pereiraPas encore d'évaluation
- Trabalho de BiologiaDocument14 pagesTrabalho de Biologiaisalves13100% (1)
- PROTOZOARIOSDocument11 pagesPROTOZOARIOSDavid Oliveira MarcussoPas encore d'évaluation
- Modulo 1 - Introdução A AnatomiaDocument26 pagesModulo 1 - Introdução A AnatomialeealencarPas encore d'évaluation
- FungosDocument14 pagesFungosEdson VicentePas encore d'évaluation
- SIDA: Uma visão geral da epidemiaDocument15 pagesSIDA: Uma visão geral da epidemiamabro abreuPas encore d'évaluation
- Reino Protista PDFDocument39 pagesReino Protista PDFEliel Mário0% (1)
- ProtozoáriosDocument3 pagesProtozoáriosNayaraPas encore d'évaluation
- Importância dos Fungos MédicosDocument36 pagesImportância dos Fungos Médicoskellen_paganiPas encore d'évaluation
- Os principais grupos de artrópodes: características, classificação e importânciaDocument13 pagesOs principais grupos de artrópodes: características, classificação e importânciaMunlhena ArturPas encore d'évaluation
- Ciclo Reprodutivo Da MulherDocument2 pagesCiclo Reprodutivo Da MulherLiliane CostaPas encore d'évaluation
- Introdução A Botanica SistemáticaDocument7 pagesIntrodução A Botanica SistemáticaIsabel Matunha MatunhaPas encore d'évaluation
- Apresentação BS Do 2 Grupo Myxobionta - PPTX 2023Document19 pagesApresentação BS Do 2 Grupo Myxobionta - PPTX 2023George Da JosinaPas encore d'évaluation
- Meu Slide FloraçãoDocument5 pagesMeu Slide FloraçãoLucas LemosPas encore d'évaluation
- Apostila de Zoologia GeralDocument30 pagesApostila de Zoologia GeralJulia MaiaPas encore d'évaluation
- Adaptações anatômicas das plantas à secaDocument7 pagesAdaptações anatômicas das plantas à secaLIANDRA BRASIL PIRESPas encore d'évaluation
- Reino PlntaeDocument37 pagesReino PlntaeMário Francisco JúniorPas encore d'évaluation
- Livro de SociologiaDocument39 pagesLivro de SociologiaJoão Vitor Alves100% (1)
- Classificação Dos Seres Vivos PDFDocument3 pagesClassificação Dos Seres Vivos PDFGiseleDuvalPas encore d'évaluation
- A história da descoberta da AidsDocument7 pagesA história da descoberta da AidsJuliana Mendonça SilvaPas encore d'évaluation
- Lei Do EfeitoDocument17 pagesLei Do EfeitoSara FernandesPas encore d'évaluation
- Fatores Limitantes e Regulatc3b3riosDocument42 pagesFatores Limitantes e Regulatc3b3riosHenrique VianaPas encore d'évaluation
- Apostila de Microbiologia 01Document12 pagesApostila de Microbiologia 01ThalitaMoraesPas encore d'évaluation
- Introdução de Linguas Bantu 2023Document14 pagesIntrodução de Linguas Bantu 2023ArquitecturaPas encore d'évaluation
- Biologia Das Algas e Plantas PDFDocument49 pagesBiologia Das Algas e Plantas PDFFCiências0% (1)
- Acessibilidade em escolas e a inclusão de alunos com deficiência motoraDocument8 pagesAcessibilidade em escolas e a inclusão de alunos com deficiência motoraEdmilson Estevão AzevedoPas encore d'évaluation
- Ciclo de Vida Dos FungosDocument21 pagesCiclo de Vida Dos FungosTayanni LucenaPas encore d'évaluation
- 8.1 Sistemas de ClassificacaoDocument3 pages8.1 Sistemas de ClassificacaosemduvidasPas encore d'évaluation
- Classificação taxonômica: categorias e reinos dos seres vivosDocument23 pagesClassificação taxonômica: categorias e reinos dos seres vivosizsharyPas encore d'évaluation
- Malnutrição grave e suas característicasDocument31 pagesMalnutrição grave e suas característicasSergio Bachir AssanePas encore d'évaluation
- O Conceito de Homologia - Darwin e A BiologiaDocument18 pagesO Conceito de Homologia - Darwin e A BiologiaAlexandre Grande MadredeusPas encore d'évaluation
- Ascaridíase - Parasitismo pelo Ascaris lumbricoidesDocument45 pagesAscaridíase - Parasitismo pelo Ascaris lumbricoidesCiasites Sites100% (1)
- Lycopodiophyta: características das licófitasDocument31 pagesLycopodiophyta: características das licófitasBianca PintoPas encore d'évaluation
- Origem Do HivDocument11 pagesOrigem Do HivMonica Ali SelemanePas encore d'évaluation
- Trabalho Sobre HIVDocument14 pagesTrabalho Sobre HIVdivano 044Pas encore d'évaluation
- Os principais tipos de reprodução assexuada em animaisDocument33 pagesOs principais tipos de reprodução assexuada em animaisTainá SilvaPas encore d'évaluation
- AlgasDocument9 pagesAlgasleoviagemPas encore d'évaluation
- Transgênicos Animal e VegetalDocument32 pagesTransgênicos Animal e VegetalAlessandra VasconcellosPas encore d'évaluation
- Fotomorfogenese PDFDocument30 pagesFotomorfogenese PDFRafael da RochaPas encore d'évaluation
- Maza HelpDocument7 pagesMaza HelpSalvador ArturPas encore d'évaluation
- Zoologia Dos InvertebradosDocument70 pagesZoologia Dos InvertebradosJalison Rêgo0% (1)
- Evolução das licófitasDocument23 pagesEvolução das licófitasGleiciane SantosPas encore d'évaluation
- As principais partes da plantaDocument2 pagesAs principais partes da plantaPatrícia Ramos100% (1)
- Coevolução inseto-plantaDocument8 pagesCoevolução inseto-plantaAna Paula MendesPas encore d'évaluation
- Aula 2 Regras de Nomenclatura Zoologica 2015 1Document27 pagesAula 2 Regras de Nomenclatura Zoologica 2015 1Tiago Silva AlmeidaPas encore d'évaluation
- Artrópodes: Características e grupos principaisDocument2 pagesArtrópodes: Características e grupos principaisGabriela AlvesPas encore d'évaluation
- Ensino GeografiaDocument5 pagesEnsino GeografiaDanilo Tadeu FélixPas encore d'évaluation
- A Importância Do Sistema DigestivoDocument23 pagesA Importância Do Sistema DigestivoRitaPinto37Pas encore d'évaluation
- I - Definição de Filosofia, Breve Contextualização e DisciplinasDocument51 pagesI - Definição de Filosofia, Breve Contextualização e DisciplinasMariaPas encore d'évaluation
- Vírus Fungos e Bacterias...Document4 pagesVírus Fungos e Bacterias...Patrícia TenórioPas encore d'évaluation
- Aula 3 ProtozoáriosDocument43 pagesAula 3 ProtozoáriosAriana AlvesPas encore d'évaluation
- TeologiaDocument11 pagesTeologiaClelioPas encore d'évaluation
- Teologia Narrativa - O Evangelho No Idioma Da Pós-ModernidadeDocument15 pagesTeologia Narrativa - O Evangelho No Idioma Da Pós-ModernidadeAlexandre GonçalvesPas encore d'évaluation
- Problemas AmbientaisDocument13 pagesProblemas AmbientaisMário António Nhanombe100% (1)
- Animais invertebrados: introdução aos principais gruposDocument11 pagesAnimais invertebrados: introdução aos principais gruposRibex JosePas encore d'évaluation
- Idade Adulta ResumoDocument3 pagesIdade Adulta ResumoMateus FélixPas encore d'évaluation
- Hormônios masculinos fisiologiaDocument8 pagesHormônios masculinos fisiologiaLuanna MotterPas encore d'évaluation
- Sistema EndócrinoDocument52 pagesSistema EndócrinocamilaamenezessalustianoPas encore d'évaluation
- Introdução à Endocrinologia e seus Principais HormôniosDocument18 pagesIntrodução à Endocrinologia e seus Principais HormôniosWaldman Santos100% (1)
- Sistema Endócrino e seus Principais HormôniosDocument9 pagesSistema Endócrino e seus Principais HormôniosFrancisco Moisés VicentePas encore d'évaluation
- Assinatura eletrônicaDocument1 pageAssinatura eletrônicaThiago Henrique OliveiraPas encore d'évaluation
- DOC-20231027-WA0013Document12 pagesDOC-20231027-WA0013Thiago Henrique OliveiraPas encore d'évaluation
- Art RiteDocument2 pagesArt RiteSoninha MotaPas encore d'évaluation
- Biofísica Do Sistema CirculatórioDocument3 pagesBiofísica Do Sistema CirculatórioLuciana Cassia50% (2)
- Gráficos Fiboh 2Document5 pagesGráficos Fiboh 2Thiago Henrique OliveiraPas encore d'évaluation
- Texto - Tecnicas Gerais de EnfermagemDocument81 pagesTexto - Tecnicas Gerais de EnfermagemFernando CarvalhoPas encore d'évaluation
- Atestado ClarísiaDocument1 pageAtestado ClarísiaThiago Henrique OliveiraPas encore d'évaluation
- TI Dispositivo pediátrico não invasivo.docx (1)Document7 pagesTI Dispositivo pediátrico não invasivo.docx (1)Thiago Henrique OliveiraPas encore d'évaluation
- 1) Questões Sistema Nervoso 1Document1 page1) Questões Sistema Nervoso 1Thiago Henrique OliveiraPas encore d'évaluation
- A história da Vigilância Sanitária e sua importância para a saúde públicaDocument58 pagesA história da Vigilância Sanitária e sua importância para a saúde públicaJujubahiaPas encore d'évaluation
- Avaliação gruposDocument3 pagesAvaliação gruposThiago Henrique OliveiraPas encore d'évaluation
- GANHO DE PESO EM RATAS PÓS Apresentação Do Dia 20 de OutubroDocument6 pagesGANHO DE PESO EM RATAS PÓS Apresentação Do Dia 20 de OutubroThiago Henrique OliveiraPas encore d'évaluation
- Cópia de HDA PDFDocument12 pagesCópia de HDA PDFThiago Henrique OliveiraPas encore d'évaluation
- Terminologias médicasDocument15 pagesTerminologias médicasThiago Henrique OliveiraPas encore d'évaluation
- SAVC Síndromes Coronárias Agudas e AVCDocument8 pagesSAVC Síndromes Coronárias Agudas e AVCThiago Henrique OliveiraPas encore d'évaluation
- Suporte Avançado de Vida Cardiovascular Manual Do ProfissionalDocument166 pagesSuporte Avançado de Vida Cardiovascular Manual Do ProfissionalThiago Henrique Oliveira100% (6)
- SAVC PCR, Arritmias e Seu TratamentoDocument6 pagesSAVC PCR, Arritmias e Seu TratamentoThiago Henrique OliveiraPas encore d'évaluation
- Cópia de HDA PDFDocument12 pagesCópia de HDA PDFThiago Henrique OliveiraPas encore d'évaluation
- Avaliação Do Ensino de Relacionamento EnfermeiroDocument2 pagesAvaliação Do Ensino de Relacionamento EnfermeiroThiago Henrique OliveiraPas encore d'évaluation
- Clínica+pDocument11 pagesClínica+pThiago Henrique OliveiraPas encore d'évaluation
- Informações para Utilização Da Taxa de BancadaDocument3 pagesInformações para Utilização Da Taxa de BancadaWalter Andrés Páez GaviriaPas encore d'évaluation
- A empatia no cuidado terapêuticoDocument3 pagesA empatia no cuidado terapêuticoAndreia CabecinhasPas encore d'évaluation
- Amor de Balada em DDocument2 pagesAmor de Balada em DThiago Henrique OliveiraPas encore d'évaluation
- Postpartum Depression - A Literature Revision Depresión Puerperal - Uma Revisión de La LiteraturaDocument8 pagesPostpartum Depression - A Literature Revision Depresión Puerperal - Uma Revisión de La LiteraturaThiago Henrique OliveiraPas encore d'évaluation
- Análise de texto sobre gestão de projetosDocument2 pagesAnálise de texto sobre gestão de projetosThiago Henrique OliveiraPas encore d'évaluation
- Gradação e Estadiamento Histopatológico de TumoresDocument25 pagesGradação e Estadiamento Histopatológico de TumoresThiago Henrique OliveiraPas encore d'évaluation
- Taxa de Bancada: Regras para uso de recursosDocument2 pagesTaxa de Bancada: Regras para uso de recursosThiago Henrique OliveiraPas encore d'évaluation
- Cifra Club - Chico César - Reis Do AgronegócioDocument5 pagesCifra Club - Chico César - Reis Do AgronegócioThiago Henrique OliveiraPas encore d'évaluation
- Cifra Club - Almir Sater - ChalanaDocument2 pagesCifra Club - Almir Sater - ChalanaThiago Henrique Oliveira100% (1)
- Cifra Club - Alceu Valença - Sete Desejos PDFDocument2 pagesCifra Club - Alceu Valença - Sete Desejos PDFThiago Henrique OliveiraPas encore d'évaluation
- Sistema circulatório humanoDocument11 pagesSistema circulatório humanoBárbara VidalPas encore d'évaluation
- Sistemas sanguíneos e grupos sanguíneosDocument1 pageSistemas sanguíneos e grupos sanguíneosAugusto CésarPas encore d'évaluation
- Apostila Pré-Vestibular - Todas As MatériasDocument566 pagesApostila Pré-Vestibular - Todas As Matériaspedroluisilva100% (3)
- Anatomia OlhoDocument3 pagesAnatomia OlhoDoris Stela LopesPas encore d'évaluation
- Procedimentos complementares plano odontológico superiorDocument10 pagesProcedimentos complementares plano odontológico superiorPAULO CESAR PADILHAPas encore d'évaluation
- Sistema Nervoso: Funções, Estrutura e OrganizaçãoDocument35 pagesSistema Nervoso: Funções, Estrutura e OrganizaçãoAmanda VianaPas encore d'évaluation
- 8 Ano CDocument3 pages8 Ano CLeon SantosPas encore d'évaluation
- Pele e AnexosDocument6 pagesPele e AnexosKamilla LFPas encore d'évaluation
- Procedimentos Tabela CISVALE 2015 OkDocument19 pagesProcedimentos Tabela CISVALE 2015 OkJackson LiraPas encore d'évaluation
- Banco de Sangue: Tipagem ABO, Rh e PráticaDocument32 pagesBanco de Sangue: Tipagem ABO, Rh e PráticaAline HúngaroPas encore d'évaluation
- Manual de NecropsiaDocument14 pagesManual de NecropsiaLouis MiyauchiPas encore d'évaluation
- Mucosa bucalDocument6 pagesMucosa bucalMabePas encore d'évaluation
- Aula 14 Funcao Glomérulo-Tubular. Depuracao PlasmáticaDocument61 pagesAula 14 Funcao Glomérulo-Tubular. Depuracao PlasmáticaJanuário Virgílio DabaliPas encore d'évaluation
- Protocolo Angio RMDocument102 pagesProtocolo Angio RMthiagoorib.21Pas encore d'évaluation
- Reações Transfusionais Tardias - USPDocument29 pagesReações Transfusionais Tardias - USPLuiza Pimenta Rochael100% (1)
- Estudo Dirigido Sistema CirculatórioDocument10 pagesEstudo Dirigido Sistema CirculatóriokievnnyPas encore d'évaluation
- GRUPOS SANGUÍNEOS E HEREDITARIEDADEDocument3 pagesGRUPOS SANGUÍNEOS E HEREDITARIEDADECarlos Ramon Andrade de OliveiraPas encore d'évaluation
- O Sistema LímbicoDocument14 pagesO Sistema LímbicoAngelita ValcanaiaPas encore d'évaluation
- Biofísica Músculo ContraçãoDocument30 pagesBiofísica Músculo ContraçãoJô Joserlane Xavier100% (3)
- Efeitos Fisiológicos Da ImersãoDocument7 pagesEfeitos Fisiológicos Da Imersãojaqueline.santosPas encore d'évaluation
- Anomalias dentárias radiografiaDocument7 pagesAnomalias dentárias radiografiaLuana Souza100% (1)
- Sequência Didática - Sistema DigestórioDocument24 pagesSequência Didática - Sistema DigestórioLi Camila RosaPas encore d'évaluation
- Enteropatia Perdedora de ProteinasDocument3 pagesEnteropatia Perdedora de ProteinasThiago de PaulaPas encore d'évaluation
- Aula+ +salivaDocument33 pagesAula+ +salivaGabriel EstevesPas encore d'évaluation
- Trombose Venosa e ArterialDocument44 pagesTrombose Venosa e ArterialVanessa SuanaPas encore d'évaluation
- Estômago CinthiaDocument59 pagesEstômago CinthiaWanniely KussPas encore d'évaluation
- Funções reprodutivas e hormonais masculinasDocument9 pagesFunções reprodutivas e hormonais masculinasRicardo Gomes Fernandes0% (1)
- Períodos da hematopoese e diferenciação celularDocument14 pagesPeríodos da hematopoese e diferenciação celulardanimb100% (1)
- Apostila Maio 2014Document15 pagesApostila Maio 2014Aleksander Milunovic100% (1)
- PRC e Hemograma de Joaquim TeixeiraDocument2 pagesPRC e Hemograma de Joaquim TeixeiraJulianna Ferreira De AraujoPas encore d'évaluation