Vous aimerez peut-être aussi
- Preinforme Fisio 2.0Document6 pagesPreinforme Fisio 2.0juako91100% (1)
- Fisiología Del GustoDocument2 pagesFisiología Del Gustozoda100% (1)
- Trabajo Completo de La LenguaDocument30 pagesTrabajo Completo de La LenguaAnnlivertyPas encore d'évaluation
- Trabajo Odontología ComunitariaDocument4 pagesTrabajo Odontología ComunitariaJose Elias Pérez Velásquez100% (1)
- Examen Parcial Nervio Trigémino y FacialDocument3 pagesExamen Parcial Nervio Trigémino y FacialAldahir100% (1)
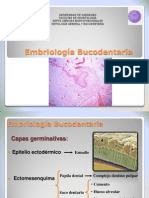
- Embriologia BucodentalDocument19 pagesEmbriologia BucodentalLorena MarinPas encore d'évaluation
- Componentes de La Reacción InflamatoriaDocument3 pagesComponentes de La Reacción InflamatoriaGameXPas encore d'évaluation
- Guia de Laboratorio Virtual y Casero Membrana CelularDocument13 pagesGuia de Laboratorio Virtual y Casero Membrana CelularOmar JallerPas encore d'évaluation
- La Interacción Paciente-Dentista, A Partir Del Significado Psicológico de La Boca PDFDocument7 pagesLa Interacción Paciente-Dentista, A Partir Del Significado Psicológico de La Boca PDFCamilo De La JaraPas encore d'évaluation
- Bases Biologicas Del PsiquismoDocument5 pagesBases Biologicas Del PsiquismoSeJo Hermoza CarrascoPas encore d'évaluation
- DegluciónDocument16 pagesDegluciónKekita Ita-itaPas encore d'évaluation
- INMUNOLOGÍA - PPTXDocument31 pagesINMUNOLOGÍA - PPTXRafael ValenzuelaPas encore d'évaluation
- Teoría CelularDocument9 pagesTeoría CelularKaren Alejandra Madrigal GonzalezPas encore d'évaluation
- Informe de Hueso AlveolarDocument14 pagesInforme de Hueso Alveolarzohan1889Pas encore d'évaluation
- Fisiologia Del Sistema EstomatognaticoDocument39 pagesFisiologia Del Sistema EstomatognaticoSheyla Espinoza Rios100% (1)
- Funciones Del Sistema MasticatorioDocument34 pagesFunciones Del Sistema MasticatorioJeon JungkookPas encore d'évaluation
- Bioquimica de La SalivaDocument18 pagesBioquimica de La SalivaMario Torres FloresPas encore d'évaluation
- Análisis de Documentos Sobre Sarro Dental y Calculo Dental-Velasco RafaelDocument11 pagesAnálisis de Documentos Sobre Sarro Dental y Calculo Dental-Velasco RafaelRafael VelascoPas encore d'évaluation
- Textoguadetrabajosprcticosubo 111122124056 Phpapp02 PDFDocument91 pagesTextoguadetrabajosprcticosubo 111122124056 Phpapp02 PDFElena CorralesPas encore d'évaluation
- Fisiología Del Sistema NerviosoDocument6 pagesFisiología Del Sistema NerviosoMauge AyalaPas encore d'évaluation
- Embriología Del OídoDocument28 pagesEmbriología Del OídoJOSE LEANDRO VASQUEZ IDROGOPas encore d'évaluation
- Semiologia Del Nervio TrigéminoDocument5 pagesSemiologia Del Nervio TrigéminoEinar Valdez EscobarPas encore d'évaluation
- Cuadro Comparativo Nivel Tisular EDIT 1Document14 pagesCuadro Comparativo Nivel Tisular EDIT 1katerin colindresPas encore d'évaluation
- CavidadesDocument33 pagesCavidadesBzaPas encore d'évaluation
- Especializaciones de MembranaDocument6 pagesEspecializaciones de MembranaNany HeddrichPas encore d'évaluation
- Mucosa BucalDocument23 pagesMucosa BucalDeniss Rios Arostegui100% (1)
- Componentes Del Sistema EstomatognaticoDocument7 pagesComponentes Del Sistema EstomatognaticoAnai Meza100% (1)
- Biología III SISTEMA ÓSEODocument4 pagesBiología III SISTEMA ÓSEOIgnacio Madrid CanalesPas encore d'évaluation
- Regiones Profundas de La CaraDocument5 pagesRegiones Profundas de La CaraHaroldChagüezaPas encore d'évaluation
- Histología de La Cavidad BucalDocument5 pagesHistología de La Cavidad BucalAli DavengerPas encore d'évaluation
- Características Histológicas de La Mucosa BucalDocument1 pageCaracterísticas Histológicas de La Mucosa BucalLeonardo Vega100% (1)
- Embriología DentaríaDocument3 pagesEmbriología DentaríaThania Milagros SANTOS PARI100% (1)
- Pulpa DentalDocument4 pagesPulpa DentalMaria SalazarPas encore d'évaluation
- Potencial de Acción e Impulso NerviosoDocument35 pagesPotencial de Acción e Impulso NerviosoFranciscoCerdaLealPas encore d'évaluation
- Mecanismos Neuromusculares Periféricos o SensorialesDocument12 pagesMecanismos Neuromusculares Periféricos o SensorialesSara CardeñoPas encore d'évaluation
- RITMO CICARDIANO-informeDocument2 pagesRITMO CICARDIANO-informekatherin PC100% (1)
- Componentes Del OídoDocument6 pagesComponentes Del OídoJovanuzqa Jatsuky McPas encore d'évaluation
- Cap 48 GuytonDocument4 pagesCap 48 GuytonCinthia LemusPas encore d'évaluation
- Clase 1 Hitologia Sistema Digestivo 2016Document44 pagesClase 1 Hitologia Sistema Digestivo 2016Patricia CastellanosPas encore d'évaluation
- Bases Sociales Del PsquismoDocument14 pagesBases Sociales Del PsquismoLucero LimacoPas encore d'évaluation
- Presentacion Acerca de La Pulpa Dental-Grupo BDocument14 pagesPresentacion Acerca de La Pulpa Dental-Grupo BTania JudithPas encore d'évaluation
- CICLO CELULAR y Mecanismos de RegulaciónDocument29 pagesCICLO CELULAR y Mecanismos de RegulaciónSalvador ResendizPas encore d'évaluation
- Macroanatomia y Microanatomia Del PeriodontDocument97 pagesMacroanatomia y Microanatomia Del PeriodontJuan Carlos Cori Soria75% (4)
- Morfogénesis Del Órgano DentarioDocument5 pagesMorfogénesis Del Órgano DentarioOscar Luis100% (1)
- Cuestionario Desarrollo de La Oclusion y La DenticionDocument1 pageCuestionario Desarrollo de La Oclusion y La DenticionLucero Iza TorresPas encore d'évaluation
- Tema 12. La Célula. Componentes Físicos y Químicos. Concepto Del Tejido.Document77 pagesTema 12. La Célula. Componentes Físicos y Químicos. Concepto Del Tejido.Loly Caamaño Lata100% (1)
- Mucosa BucalDocument42 pagesMucosa BucalAnonymous 1tIQ72Pas encore d'évaluation
- Aparato Genital Masculino Guias13Document9 pagesAparato Genital Masculino Guias13Luis GallinaPas encore d'évaluation
- Expo Cirugia AnestesiaDocument36 pagesExpo Cirugia AnestesiaIlich Garay100% (2)
- Sistema EndocrinoDocument39 pagesSistema EndocrinoNatalia JaramilloPas encore d'évaluation
- Fisiologia Del OidoDocument20 pagesFisiologia Del OidoIvette GonzálezPas encore d'évaluation
- Señalización Celular 2018Document80 pagesSeñalización Celular 2018Jafet RíosPas encore d'évaluation
- Caries DentalDocument30 pagesCaries DentalMark L. Sandoval VegaPas encore d'évaluation
- Arcadas DentariasDocument5 pagesArcadas DentariasElizabethPas encore d'évaluation
- Intro Historia de Los AnestésicosDocument12 pagesIntro Historia de Los AnestésicosGonzalo Paredes GonzalesPas encore d'évaluation
- Morfogénesis Del Órgano DentarioDocument4 pagesMorfogénesis Del Órgano DentarioAripcy MatiasPas encore d'évaluation
- Inmunidad Innata - ResumenDocument1 pageInmunidad Innata - Resumenfer byun100% (1)
- Complejo Articular TemporomandibularDocument46 pagesComplejo Articular Temporomandibularsabbath19Pas encore d'évaluation
- Receptores, EstimuloDocument2 pagesReceptores, EstimuloErik J. Roshan FloresPas encore d'évaluation
- Notas 2 ParcialDocument48 pagesNotas 2 ParcialAlan EdmundoPas encore d'évaluation
- Antecedentes HistóricosDocument2 pagesAntecedentes HistóricosGALOXRLPas encore d'évaluation
- Prosencefalo BLDocument13 pagesProsencefalo BLbrayan_dw100% (1)
- Fisiología RespiratoriaDocument33 pagesFisiología RespiratoriaDaniel Roa BahamondesPas encore d'évaluation
- Informe - Harina de SangreDocument19 pagesInforme - Harina de SangreEliana Rodriguez100% (1)
- Medula Espinal Anatomofisiologia AsgDocument18 pagesMedula Espinal Anatomofisiologia AsgHumberto CarvajalPas encore d'évaluation
- Especies en Los Bofedales de HUaytireDocument22 pagesEspecies en Los Bofedales de HUaytirenosekeponerploxPas encore d'évaluation
- Alimentacion General Especializada Mascotas PDFDocument50 pagesAlimentacion General Especializada Mascotas PDFSebastian Lopez ArangoPas encore d'évaluation
- Elaboración de QuesosDocument15 pagesElaboración de Quesosbetty.lenis100% (1)
- Cuidado de Higiene Oral en Niños de 1 A 6 AñosDocument1 pageCuidado de Higiene Oral en Niños de 1 A 6 AñosLINA ORTIZPas encore d'évaluation
- Triangulos Del CuelloDocument9 pagesTriangulos Del CuelloErick Montenegro100% (1)
- ImagoDocument202 pagesImagoCharly RiañoPas encore d'évaluation
- Mendiola Rivera Mauricio - PRACTICA-17 (Autoguardado)Document5 pagesMendiola Rivera Mauricio - PRACTICA-17 (Autoguardado)Mauricio Mendiola RiveraPas encore d'évaluation
- Competencias 16 Agosto 2013 Definitivo 2Document53 pagesCompetencias 16 Agosto 2013 Definitivo 2YENNY ALEJANDRA NEGRETE SANCHEZPas encore d'évaluation
- Informe de Reductasa de LecheDocument11 pagesInforme de Reductasa de LecheFrancel ObregoPas encore d'évaluation
- Alveoloplastia y Regulación de Los Rebordes AlveolaresDocument18 pagesAlveoloplastia y Regulación de Los Rebordes AlveolaresAdriana Escolar Ruiz75% (4)
- Cambios Fisiológicos en El EmbarazoDocument24 pagesCambios Fisiológicos en El Embarazocarmen gonzalez100% (3)
- Resumen Cadena EpidemiologicaDocument3 pagesResumen Cadena EpidemiologicaLeykyn Vargas50% (2)
- Sasquatch Enigma AntropológicoDocument177 pagesSasquatch Enigma AntropológicoSWAT666Pas encore d'évaluation
- Taller OctavoDocument1 pageTaller OctavoTsukiHikariAnjera100% (3)
- Índice de IHOSDocument11 pagesÍndice de IHOSIsaac Og100% (2)
- Adaptaciones de Animales y Plantas para La Conservación Del AguaDocument7 pagesAdaptaciones de Animales y Plantas para La Conservación Del Aguaep00124100% (1)
- Topicos Parasitologia Modulo IVaDocument5 pagesTopicos Parasitologia Modulo IVaJulio YarPas encore d'évaluation
- Ebbo Opon Miguel FeblesDocument21 pagesEbbo Opon Miguel Feblesoowonrin100% (2)
- Fosas Del CraneoDocument67 pagesFosas Del CraneoMedSchoolStuffPas encore d'évaluation
- Monografía Paleolítico y NeolíticoDocument4 pagesMonografía Paleolítico y NeolíticoSebastian Agosta50% (2)
- Dominio Linguístico - Tarea 3Document22 pagesDominio Linguístico - Tarea 3jaime figueroa40% (5)
- Anticuerpos Anticelulas ParietalesDocument2 pagesAnticuerpos Anticelulas ParietalesCrysthPas encore d'évaluation
- Prueba Lenguaje Gregor y La Profesia Del Gris Septimo AñoDocument3 pagesPrueba Lenguaje Gregor y La Profesia Del Gris Septimo AñoJuan Carlos Barriga PeñaPas encore d'évaluation
- Textos Narrativos 3Document25 pagesTextos Narrativos 3Jhon Coronel JorgePas encore d'évaluation
- Pork Pig Producers SpainDocument16 pagesPork Pig Producers SpainGarrettMcGuckiangmail.comPas encore d'évaluation