Vous aimerez peut-être aussi
- DengueDocument7 pagesDengueeddumedinaPas encore d'évaluation
- Senasa EntrevistaDocument7 pagesSenasa EntrevistaeddumedinaPas encore d'évaluation
- 09464858Document1 page09464858eddumedinaPas encore d'évaluation
- Opinión Documentada S1-T6Document2 pagesOpinión Documentada S1-T6eddumedinaPas encore d'évaluation
- El AjedrezDocument1 pageEl AjedrezeddumedinaPas encore d'évaluation
- Pulmones 131220170341 Phpapp01Document10 pagesPulmones 131220170341 Phpapp01William OscarPas encore d'évaluation
- Seminario 14Document44 pagesSeminario 14eddumedinaPas encore d'évaluation
- Diapos FinalDocument35 pagesDiapos FinaleddumedinaPas encore d'évaluation
- Declaracion Jurada ADocument2 pagesDeclaracion Jurada AJose Miguel Beingolea ValenzaPas encore d'évaluation
- Fagocitosis Tipos y ClasificacionDocument4 pagesFagocitosis Tipos y ClasificacioneddumedinaPas encore d'évaluation
- Hormona Del Crecimiento Ejerce Varios Efectos MetabólicosDocument7 pagesHormona Del Crecimiento Ejerce Varios Efectos MetabólicoseddumedinaPas encore d'évaluation
- Gastocardiacoretornovenosoysuregulacion 141202124828 Conversion Gate01Document127 pagesGastocardiacoretornovenosoysuregulacion 141202124828 Conversion Gate01eddumedinaPas encore d'évaluation
- Caso Clinico 4Document1 pageCaso Clinico 4eddumedinaPas encore d'évaluation
- Fisio EkgDocument9 pagesFisio EkgeddumedinaPas encore d'évaluation
- Patrones de Velas JaponesasDocument3 pagesPatrones de Velas JaponesaseddumedinaPas encore d'évaluation
- Ganglios y NeutrofilosDocument4 pagesGanglios y NeutrofiloseddumedinaPas encore d'évaluation
- Patrones de Velas JaponesasDocument9 pagesPatrones de Velas JaponesasLuis RealPas encore d'évaluation
- ElisaDocument3 pagesElisaeddumedinaPas encore d'évaluation
- Funciones reproductoras masculinas y hormonasDocument12 pagesFunciones reproductoras masculinas y hormonaseddumedinaPas encore d'évaluation
- Retinopatía Hipertensiva y Diabética en Paciente de 80 AñosDocument1 pageRetinopatía Hipertensiva y Diabética en Paciente de 80 AñoseddumedinaPas encore d'évaluation
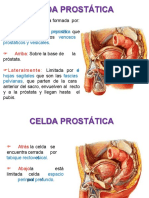
- Celda ProstaticaDocument7 pagesCelda ProstaticaeddumedinaPas encore d'évaluation
- Fisio Ultima ParteDocument12 pagesFisio Ultima ParteeddumedinaPas encore d'évaluation
- GHC ElisaDocument7 pagesGHC ElisaeddumedinaPas encore d'évaluation
- Dolor FisioDocument14 pagesDolor FisioeddumedinaPas encore d'évaluation
- Anatomia EdduDocument7 pagesAnatomia EddueddumedinaPas encore d'évaluation
- Universidad Privada Antenor OrregoDocument1 pageUniversidad Privada Antenor OrregoeddumedinaPas encore d'évaluation
- Banco de Inmuno SustiDocument83 pagesBanco de Inmuno SustieddumedinaPas encore d'évaluation
- Mustard de Spain Sobre NeurologiaDocument13 pagesMustard de Spain Sobre NeurologiaJenifferPas encore d'évaluation
- LinksDocument1 pageLinkseddumedinaPas encore d'évaluation
- Registros IntermaxilaresDocument35 pagesRegistros IntermaxilaresRay Ruz Dominguez67% (3)
- Ficha Evaluación Fonoaudiológica OFASDocument1 pageFicha Evaluación Fonoaudiológica OFASValeria Campos CallecPas encore d'évaluation
- Meninges, LCR, Irrigación y Drenaje PDFDocument46 pagesMeninges, LCR, Irrigación y Drenaje PDFFernandoAlexanderRamirezZamoraPas encore d'évaluation
- Fisiopatologia de La InflamacionDocument8 pagesFisiopatologia de La InflamacionCarla100% (1)
- Células eucariotas y procariotasDocument17 pagesCélulas eucariotas y procariotasRomina Dome100% (1)
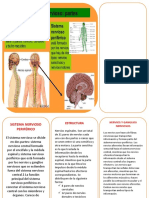
- Sistema Nervioso PeriféricoDocument2 pagesSistema Nervioso PeriféricoRichard HuancaPas encore d'évaluation
- JUSTIFICACIÓNDocument19 pagesJUSTIFICACIÓNDavid100% (1)
- El Cerebro en Desarrollo 2012Document38 pagesEl Cerebro en Desarrollo 2012Gerard Fisa BiscarriPas encore d'évaluation
- EndocrinologíaDocument21 pagesEndocrinologíaJoan ZambranoPas encore d'évaluation
- ArticulacionesDocument52 pagesArticulacionesMax Perz PerzPas encore d'évaluation
- Cuestionario Sobre División Topográfica Del Abdomen - Attempt ReviewDocument3 pagesCuestionario Sobre División Topográfica Del Abdomen - Attempt ReviewKatrina AlcantaraPas encore d'évaluation
- Pared Abdominal AnteriorDocument2 pagesPared Abdominal AnteriorIsra Waafles' GomezPas encore d'évaluation
- DiverticulosisDocument50 pagesDiverticulosisPepeQPas encore d'évaluation
- HormonasDocument7 pagesHormonasRoberto Cuevas100% (1)
- AyudinDocument13 pagesAyudinYocelin FernándezPas encore d'évaluation
- Endocrino PreguntasDocument5 pagesEndocrino PreguntasNataly Jorquera CopaivaPas encore d'évaluation
- Manual MSD DocumentDocument5 pagesManual MSD Documentyohanaegm6Pas encore d'évaluation
- Manchas de Sangre PDFDocument6 pagesManchas de Sangre PDFbelisario66Pas encore d'évaluation
- Anatomía II - CuestionarioDocument8 pagesAnatomía II - CuestionarioMariuxi Elizabeth PereroPas encore d'évaluation
- Reporte Práctica 11Document23 pagesReporte Práctica 11Meli CastilloPas encore d'évaluation
- Sistema Nervioso Central: IntegrantesDocument27 pagesSistema Nervioso Central: IntegrantesIsabel Alejandra Romero AlvaPas encore d'évaluation
- Caso Clinico HermafroditaDocument7 pagesCaso Clinico HermafroditajorgecarpenaPas encore d'évaluation
- Barrera Hematoencefalica.Document13 pagesBarrera Hematoencefalica.Alfonso Quispe CopaPas encore d'évaluation
- Desarrollo de Maxila y MandibulaDocument23 pagesDesarrollo de Maxila y MandibulaXavier Alexander100% (7)
- Fisiologia Ciclo Sexual FemeninoDocument19 pagesFisiologia Ciclo Sexual FemeninoAlmendra alba MontenegroPas encore d'évaluation
- Sistema ExcretorDocument10 pagesSistema ExcretorYAZMIN CORREDOR CORZOPas encore d'évaluation
- Migracion Erratica PDFDocument6 pagesMigracion Erratica PDFElenita RojasPas encore d'évaluation
- Órganos Del Sistema InmuneDocument24 pagesÓrganos Del Sistema InmuneantonietaPas encore d'évaluation
- Mediastino Jose PDFDocument114 pagesMediastino Jose PDFJasmini Meow06100% (1)