Vous aimerez peut-être aussi
- Cuadernillo Terapia de LenguajeDocument41 pagesCuadernillo Terapia de LenguajeKarim Peña Peseros100% (1)
- Caldos MicrobianosDocument38 pagesCaldos Microbianosarmandoguelfi563280% (25)
- Articulaciones y tipos de movimientoDocument47 pagesArticulaciones y tipos de movimientobhanny2Pas encore d'évaluation
- BiorremediacionDocument8 pagesBiorremediacionbrayangt100% (2)
- Caso Clínico 5 Técnica Meaw y MicroimplantesDocument41 pagesCaso Clínico 5 Técnica Meaw y MicroimplantesluisarriolaguillenPas encore d'évaluation
- Sistema de LagunajesDocument10 pagesSistema de LagunajesAnonymous WxIzg7tPas encore d'évaluation
- Anatomía y comportamiento de las serpientes enDocument13 pagesAnatomía y comportamiento de las serpientes encarlos carrilloPas encore d'évaluation
- OftalmologiaDocument97 pagesOftalmologiaUvi Cancino RamosPas encore d'évaluation
- Sistema respiratorio: anatomía y función enDocument27 pagesSistema respiratorio: anatomía y función enLady CastañedaPas encore d'évaluation
- Conducta ovinosDocument80 pagesConducta ovinospabloPas encore d'évaluation
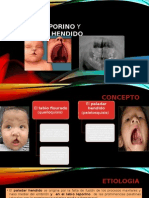
- Labio LeporinoDocument11 pagesLabio LeporinoAndres PerezPas encore d'évaluation
- CLASE DE ESQUELETO AXIAL. Clase 2 Unidad 2Document112 pagesCLASE DE ESQUELETO AXIAL. Clase 2 Unidad 2CAMILA ANDREA CONDE ARANAPas encore d'évaluation
- Anatomia Comparada Ap Reprod y UrogenitalDocument58 pagesAnatomia Comparada Ap Reprod y UrogenitalMaigualida OrbetaPas encore d'évaluation
- Aparato Digestivo ExpoDocument44 pagesAparato Digestivo ExpoRubiiXx GrPas encore d'évaluation
- Modeladores elásticos BimlerDocument66 pagesModeladores elásticos BimlerMilton David Rios Serrato67% (3)
- Guía Completa Generalidades Anatomia ULSA 2020 1Document12 pagesGuía Completa Generalidades Anatomia ULSA 2020 1Laura Sofia Avendaño Morales100% (1)
- Angiografia de Miembros InferioresDocument14 pagesAngiografia de Miembros Inferioresandres felipePas encore d'évaluation
- Manejo Del Dolor en El Paciente Crítico - Pablo Otero - Mayo2020Document141 pagesManejo Del Dolor en El Paciente Crítico - Pablo Otero - Mayo2020Dominga MuñozPas encore d'évaluation
- Neuroanatomia - Veterinaria - ContinuacionDocument5 pagesNeuroanatomia - Veterinaria - ContinuacionMarcela MoraPas encore d'évaluation
- Anatomia y Morfologia AnimalDocument4 pagesAnatomia y Morfologia AnimalJose QueralesPas encore d'évaluation
- Anatomía Del Encéfalo.Document21 pagesAnatomía Del Encéfalo.Camila Alejandra Jurado AriasPas encore d'évaluation
- 3 Aparato Digestivo NUTDocument162 pages3 Aparato Digestivo NUTareli elizabeth reynoso jimenezPas encore d'évaluation
- Anatomía y Fisiolgía de La PróstataDocument9 pagesAnatomía y Fisiolgía de La PróstataFrank Silva Fonseca0% (1)
- Ojo VacaDocument26 pagesOjo VacaLinda Valencia ValdezPas encore d'évaluation
- Anatomia de La GallinaDocument28 pagesAnatomia de La Gallinavictor alpirezPas encore d'évaluation
- Anatomía y fisiología digestiva de la cabraDocument22 pagesAnatomía y fisiología digestiva de la cabraElcoba Caballero0% (1)
- Ojo Anatomía e Histología, FinalDocument6 pagesOjo Anatomía e Histología, FinalJacob MendozaPas encore d'évaluation
- GENERALIDADES DE OSTEOLOGÍA 2020 - 2 Guia 2Document7 pagesGENERALIDADES DE OSTEOLOGÍA 2020 - 2 Guia 2Laura Sofia Avendaño MoralesPas encore d'évaluation
- Manual de Extracción de Sangre en TortugasDocument15 pagesManual de Extracción de Sangre en TortugasIsabel ContrerasPas encore d'évaluation
- Anatomia Descriptiva ComparadaDocument16 pagesAnatomia Descriptiva ComparadaJorge Espinola (Djorka)Pas encore d'évaluation
- AP DigestivoDocument76 pagesAP DigestivoSolGimenezPas encore d'évaluation
- Anatomía veterinaria tórax y dorsoDocument13 pagesAnatomía veterinaria tórax y dorsoCecilia Montoya PachasPas encore d'évaluation
- Practica de SaludDocument4 pagesPractica de SaludBernardo Resendiz GonzalezPas encore d'évaluation
- Anatomia Del Ojo Animal PDFDocument81 pagesAnatomia Del Ojo Animal PDFJorge GomezPas encore d'évaluation
- Glándulas SalivalesDocument46 pagesGlándulas SalivalesJessica0% (1)
- Fisiología Animal Comparada Del Sistema RespiratorioDocument11 pagesFisiología Animal Comparada Del Sistema RespiratorioJoseth Garcia ChirePas encore d'évaluation
- Cuadro Comparativo EndocrinoDocument7 pagesCuadro Comparativo EndocrinoTanya Rubí100% (1)
- ECOLOGIADocument3 pagesECOLOGIAVanessa JimenezPas encore d'évaluation
- Huesos del cráneo: estudio e identificaciónDocument65 pagesHuesos del cráneo: estudio e identificaciónMishelle AlexandraPas encore d'évaluation
- Ciclo EstralDocument36 pagesCiclo EstralCarlos SaucedoPas encore d'évaluation
- Diseccion TruchaDocument3 pagesDiseccion TruchaJuan Antonio RuizPas encore d'évaluation
- Origen y Clasificacion de Los Reptiles PDFDocument5 pagesOrigen y Clasificacion de Los Reptiles PDFArriane AlterPas encore d'évaluation
- 001-2014-CC-Anatomia, Introduccion PDFDocument40 pages001-2014-CC-Anatomia, Introduccion PDFgisella0% (1)
- Extinción de Los Diversos Tipos de Tortugas en El Estado de Tabasc1Document6 pagesExtinción de Los Diversos Tipos de Tortugas en El Estado de Tabasc1Karla MartinezPas encore d'évaluation
- Clase 14 Aparato Reproductor de La HembraDocument30 pagesClase 14 Aparato Reproductor de La Hembravictor quispe Castro100% (1)
- I. EsqueletoDocument30 pagesI. EsqueletoAndre Efrain Motoche LeonPas encore d'évaluation
- Clase Cestodos 2016 PDFDocument123 pagesClase Cestodos 2016 PDFCarlos EnsunchoPas encore d'évaluation
- Anatomia de AvesDocument55 pagesAnatomia de AvesGrace Katherin Muñoz GarciaPas encore d'évaluation
- Abdomen ARP - PpsDocument106 pagesAbdomen ARP - PpsErnesto RangelPas encore d'évaluation
- Pares Craneales Vii-XiiDocument35 pagesPares Craneales Vii-XiiJudith OstosPas encore d'évaluation
- Marco TeóricoDocument24 pagesMarco TeóricoJohan AndradePas encore d'évaluation
- Bazo, Bolsa de Fabricio y TimoDocument10 pagesBazo, Bolsa de Fabricio y TimoCatalina NañezPas encore d'évaluation
- URG - Articulaciones MiembrosDocument53 pagesURG - Articulaciones MiembrosXimena Olivares CastilloPas encore d'évaluation
- Cardio AvesDocument7 pagesCardio AvesLaura Castro Ramírez0% (1)
- Histologia Enfermedades Oculares Perros y GatosDocument22 pagesHistologia Enfermedades Oculares Perros y Gatosvaleria toaza leonPas encore d'évaluation
- Introduccion A La Anatomia IDocument23 pagesIntroduccion A La Anatomia IMatthew BrooksPas encore d'évaluation
- Anatomía AnimalDocument2 pagesAnatomía AnimalCamilo MéndezPas encore d'évaluation
- Las Funciones de Los Diverticulos Del RumiantesDocument6 pagesLas Funciones de Los Diverticulos Del Rumiantesedgar cundapiPas encore d'évaluation
- Fracturas en CaninosDocument1 pageFracturas en CaninosMaría Paula CuellarPas encore d'évaluation
- Enfermedad Cronica Desgastante Del VenadoDocument1 pageEnfermedad Cronica Desgastante Del VenadoMario Uziel Reyes VazquezPas encore d'évaluation
- Fotos Cuaderno AveDocument14 pagesFotos Cuaderno AveMihaela MuñozPas encore d'évaluation
- Angiología ClasesDocument23 pagesAngiología ClasesRaul Coila SalazarPas encore d'évaluation
- Conformación Pélvica en Las Diferentes EspeciesDocument4 pagesConformación Pélvica en Las Diferentes EspeciesLiz Florencia ViñabalPas encore d'évaluation
- Propedéutica VeterinariaDocument12 pagesPropedéutica VeterinariaDaniela Loranca ValdovinosPas encore d'évaluation
- Anatomía cavidad toráxica perroDocument24 pagesAnatomía cavidad toráxica perroMónica CalviñoPas encore d'évaluation
- Clase 2 Osteologia y Artrología de Craneo UssDocument54 pagesClase 2 Osteologia y Artrología de Craneo UsssebastkmPas encore d'évaluation
- Manual de prácticas de laboratorio en microbiología veterinariaD'EverandManual de prácticas de laboratorio en microbiología veterinariaPas encore d'évaluation
- Leguminosas espontaneas de posible valor forrajero en ColombiaD'EverandLeguminosas espontaneas de posible valor forrajero en ColombiaPas encore d'évaluation
- Qué Es ATP PDFDocument1 pageQué Es ATP PDFJuan José Tovar PérezPas encore d'évaluation
- 4alteracion 6541Document64 pages4alteracion 6541Joels Casanova OrtizPas encore d'évaluation
- NTC256Document9 pagesNTC256Anonymous WxIzg7tPas encore d'évaluation
- Exposicion RAEES FinalDocument8 pagesExposicion RAEES FinalAnonymous WxIzg7tPas encore d'évaluation
- Basura Electronica Otra Cara Tecnologia GreenpeaceDocument25 pagesBasura Electronica Otra Cara Tecnologia GreenpeaceCh'aska Chávez MoscosoPas encore d'évaluation
- Validacion y Verificacion de Metodos MicrobiologicosDocument6 pagesValidacion y Verificacion de Metodos MicrobiologicosAnonymous WxIzg7tPas encore d'évaluation
- Exposicion RAEES FinalDocument8 pagesExposicion RAEES FinalAnonymous WxIzg7tPas encore d'évaluation
- Copa Cerrada Pensky Martens EPA 1010Document4 pagesCopa Cerrada Pensky Martens EPA 1010mjimenezgPas encore d'évaluation
- 3m aMONIOS CUATERNARIOSDocument16 pages3m aMONIOS CUATERNARIOSAnonymous WxIzg7tPas encore d'évaluation
- Inventario LaboratorioDocument14 pagesInventario LaboratorioAnonymous WxIzg7tPas encore d'évaluation
- Determinacic3b3n de Azc3bacares Reductores Por El Mc3a9todo de FehlingDocument4 pagesDeterminacic3b3n de Azc3bacares Reductores Por El Mc3a9todo de FehlingAnonymous WxIzg7tPas encore d'évaluation
- ReporteTramite NSODocument5 pagesReporteTramite NSOAnonymous WxIzg7tPas encore d'évaluation
- Norma NFPA 704 PDFDocument3 pagesNorma NFPA 704 PDFFernando Vallejos100% (1)
- Decreto 3075 de 1997Document47 pagesDecreto 3075 de 1997Dianita RubioPas encore d'évaluation
- Manual de Procedimientos para Análisis de Aguas y Efluentes IndustrialesDocument174 pagesManual de Procedimientos para Análisis de Aguas y Efluentes IndustrialesFortunato Seijas50% (2)
- Articulo Boletin Desinfectantes y Modo de Accion en IIAADocument10 pagesArticulo Boletin Desinfectantes y Modo de Accion en IIAAJisseth ChamorroPas encore d'évaluation
- Palma de Cera AfricanaDocument22 pagesPalma de Cera AfricanaAnonymous WxIzg7tPas encore d'évaluation
- Universidad Colegio Mayor de CundinamarcaDocument2 pagesUniversidad Colegio Mayor de CundinamarcaAnonymous WxIzg7tPas encore d'évaluation
- Recomend Ac I OnesDocument8 pagesRecomend Ac I OnesChristian VargasPas encore d'évaluation
- Copas de DensidadDocument2 pagesCopas de DensidadDavid MedinaPas encore d'évaluation
- Qué Es El Balance GeneralDocument1 pageQué Es El Balance GeneralAnonymous WxIzg7tPas encore d'évaluation
- Taller Post Salida de CampoDocument8 pagesTaller Post Salida de CampoAnonymous WxIzg7tPas encore d'évaluation
- Guiaparatomademuestrasdeaguas IDEAMDocument83 pagesGuiaparatomademuestrasdeaguas IDEAMjorgem2005Pas encore d'évaluation
- Planificacion Semestral-I-2012 Ucmc Virologia ClinicaDocument20 pagesPlanificacion Semestral-I-2012 Ucmc Virologia ClinicaAnonymous WxIzg7tPas encore d'évaluation
- Cuantificación de ácidos nucleicos espectrofotométricamente (40Document4 pagesCuantificación de ácidos nucleicos espectrofotométricamente (40Anonymous WxIzg7t0% (1)
- Ciencias Basicas en OftalmologiaDocument23 pagesCiencias Basicas en OftalmologiaLorena M. FigueredoPas encore d'évaluation
- Resumen OFTALMODocument38 pagesResumen OFTALMONereaRodriguezTellePas encore d'évaluation
- Desarrollo de La Boca, Nariz y PaladarDocument20 pagesDesarrollo de La Boca, Nariz y PaladarAlvaro Vasquez Orozco100% (3)
- La VisionDocument14 pagesLa VisionjssalinasPas encore d'évaluation
- Historia Clinica Oftalmologica 3Document25 pagesHistoria Clinica Oftalmologica 3Soraya MartinezPas encore d'évaluation
- LCniñosSEODocument5 pagesLCniñosSEOLorena MendozaPas encore d'évaluation
- Monografia GlaucomaDocument38 pagesMonografia GlaucomaChinones0% (1)
- 2016 Nariz Torcida Daniels EsDocument13 pages2016 Nariz Torcida Daniels EsJuliann PedroPas encore d'évaluation
- Seno MaxilarDocument3 pagesSeno MaxilarJeferson CabreraPas encore d'évaluation
- WWW Google Com Search Ei FeiGXOj7BMWm5OUPlsS3kAY Q PDF IridoDocument3 pagesWWW Google Com Search Ei FeiGXOj7BMWm5OUPlsS3kAY Q PDF IridoserlibresPas encore d'évaluation
- Guia de Lentillas PDFDocument20 pagesGuia de Lentillas PDFLeyrePas encore d'évaluation
- El OjoDocument22 pagesEl OjoFabi CardenasPas encore d'évaluation
- Catarata FinalDocument51 pagesCatarata Finalstiven narvaezPas encore d'évaluation
- Afaquia: causas, síntomas y tratamientoDocument7 pagesAfaquia: causas, síntomas y tratamientoZayra OcambalPas encore d'évaluation
- Odontología - Detalle de CódigosDocument2 pagesOdontología - Detalle de CódigosGiu FoisPas encore d'évaluation
- ANATOMÍA Areas de CottleDocument2 pagesANATOMÍA Areas de CottleDiego HidalgoPas encore d'évaluation
- Vanessa Herrera Guerra HCDocument3 pagesVanessa Herrera Guerra HCcristobalPas encore d'évaluation
- FOTORRECEPCIÓNDocument61 pagesFOTORRECEPCIÓNGustavoPas encore d'évaluation
- El Oido y Sus PartesDocument2 pagesEl Oido y Sus PartesMayka Osuna100% (7)
- El Olfato y La Vista para Tercero de PrimariaDocument4 pagesEl Olfato y La Vista para Tercero de PrimariaMaria Lna MedinaPas encore d'évaluation
- Anatomía en El Maquillaje OjosDocument1 pageAnatomía en El Maquillaje OjosCeciliaPas encore d'évaluation
- Wuolah-Free-Temas 10-16Document28 pagesWuolah-Free-Temas 10-16Evelyn SepulvedaPas encore d'évaluation
- Embriología Del Globo OcularDocument24 pagesEmbriología Del Globo OcularAngel Gomez100% (1)
- Orto Oct16Document2 pagesOrto Oct16Amparo SolerPas encore d'évaluation
- Causas de deficiencia visual tras trauma ocularDocument2 pagesCausas de deficiencia visual tras trauma ocularrafaelPas encore d'évaluation