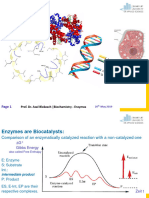
Vous aimerez peut-être aussi
- Pinealon Increases Cell Viability by Suppression of Free Radical Levels and Activating Proliferative Processes.Document8 pagesPinealon Increases Cell Viability by Suppression of Free Radical Levels and Activating Proliferative Processes.scribdPas encore d'évaluation
- Cell Free SystemDocument8 pagesCell Free SystemDuaa ShamiehPas encore d'évaluation
- Amin Et Al. - 2001 - Reactive Oxygen Species Mediate Alpha-Adrenergic Receptor-Stimulated Hypertrophy in Adult Rat Ventricular Myocytes PDFDocument9 pagesAmin Et Al. - 2001 - Reactive Oxygen Species Mediate Alpha-Adrenergic Receptor-Stimulated Hypertrophy in Adult Rat Ventricular Myocytes PDFflashjetPas encore d'évaluation
- Jamur Tiram 1Document6 pagesJamur Tiram 1Muhammad AkbarPas encore d'évaluation
- Collagen Synthesis in Cultured Human Skin Fibroblasts - Effect of AADocument5 pagesCollagen Synthesis in Cultured Human Skin Fibroblasts - Effect of AAfgonzalezPas encore d'évaluation
- SJ BJP 0706338Document8 pagesSJ BJP 0706338Rogério HitsugayaPas encore d'évaluation
- J. Biol. Chem.-1994-Jaffe-13065-8Document4 pagesJ. Biol. Chem.-1994-Jaffe-13065-8Toeii ChatpolchaiPas encore d'évaluation
- Regulación Del Metabolismo de Los Eritrocitos Por La InsulinaDocument11 pagesRegulación Del Metabolismo de Los Eritrocitos Por La InsulinaJesus D. Hernandez GuitianPas encore d'évaluation
- Factors Affecting Gluconeogenesis in The Neonatal Subhuman Primate (Macaca Mulatta)Document8 pagesFactors Affecting Gluconeogenesis in The Neonatal Subhuman Primate (Macaca Mulatta)Saleth CortezPas encore d'évaluation
- Sloane1981 - Science - CathepsinsDocument4 pagesSloane1981 - Science - CathepsinsPilar AufrastoPas encore d'évaluation
- PIIS0021925819685212Document6 pagesPIIS0021925819685212windfragPas encore d'évaluation
- CDNA Lab Report - Docx2Document4 pagesCDNA Lab Report - Docx2cxs5278100% (1)
- 2016.Zheng-Hai - Activating Nuclear Xenobiotic Receptors and Triggering ER Stress and Hepatic Cytochromes P450 Systems in Quails (Coturnix C-Coturnix) During Atrazine ExposureDocument10 pages2016.Zheng-Hai - Activating Nuclear Xenobiotic Receptors and Triggering ER Stress and Hepatic Cytochromes P450 Systems in Quails (Coturnix C-Coturnix) During Atrazine ExposureIasmin ResendePas encore d'évaluation
- Purification and Characterization of Hexokinase From Leishmania MexicanaDocument8 pagesPurification and Characterization of Hexokinase From Leishmania MexicanaWendy SailemaPas encore d'évaluation
- 016 Scas Abstracts Listed Alphabetically by Abstractspdf 19 2016 Scas AbstractsDocument25 pages016 Scas Abstracts Listed Alphabetically by Abstractspdf 19 2016 Scas AbstractsdssagPas encore d'évaluation
- Robin Solanki Botony Assignment 2Document6 pagesRobin Solanki Botony Assignment 2Saloni ChaudharyPas encore d'évaluation
- Bonney 1979Document12 pagesBonney 1979Auliana FEPas encore d'évaluation
- In Vitro Zonation and Toxicity in A Hepatocyte Bioreactor: Toxicological SciencesDocument10 pagesIn Vitro Zonation and Toxicity in A Hepatocyte Bioreactor: Toxicological SciencesRamboPas encore d'évaluation
- The Arginine Deiminase Pathway in Regulation of Arginine-Ornithine Exchange andDocument9 pagesThe Arginine Deiminase Pathway in Regulation of Arginine-Ornithine Exchange andAvishekh SinhaPas encore d'évaluation
- LP Human Pig DogDocument7 pagesLP Human Pig DogHarold Rodriguez LarrotaPas encore d'évaluation
- Increased Expression of Heparanase in Puromycin Aminonucleoside NephrosisDocument10 pagesIncreased Expression of Heparanase in Puromycin Aminonucleoside NephrosisFauziana NurhanisahPas encore d'évaluation
- Soares Et Al., 2017 Steroid Hormones Interact With Natriuretic Peptide C ToDocument8 pagesSoares Et Al., 2017 Steroid Hormones Interact With Natriuretic Peptide C ToLudimilaPas encore d'évaluation
- Saloni Chaudhary 2019659564 Botony Assignment 2Document6 pagesSaloni Chaudhary 2019659564 Botony Assignment 2Saloni ChaudharyPas encore d'évaluation
- Cancer NK Cell ActivationDocument7 pagesCancer NK Cell ActivationMazlan SijifPas encore d'évaluation
- Hildegarde Esther Allen: City of Medical AND Division OF OFDocument7 pagesHildegarde Esther Allen: City of Medical AND Division OF OFJulien Patrick CebrianPas encore d'évaluation
- TUDERMAN Et Al-1982-European Journal of BiochemistryDocument5 pagesTUDERMAN Et Al-1982-European Journal of BiochemistryAudrey Novita ArijantoPas encore d'évaluation
- Purinergic Modulation of Interleukin-1 Release From Microglial Cells Stimulated With Bacterial EndotoxinDocument4 pagesPurinergic Modulation of Interleukin-1 Release From Microglial Cells Stimulated With Bacterial EndotoxinSofi MoralesPas encore d'évaluation
- Role of Nadph:Cytochrome C Reducã-Aseand Dt-Diaphorase in The Biotransformation of Mitomycin C1Document7 pagesRole of Nadph:Cytochrome C Reducã-Aseand Dt-Diaphorase in The Biotransformation of Mitomycin C1Krys tallPas encore d'évaluation
- Beta GlukanDocument9 pagesBeta GlukanReza Dwi SaputraPas encore d'évaluation
- Stationary Phase and The Cell Cycle of Dictyostelium Discoideum in Liquid Nutrient MediumDocument11 pagesStationary Phase and The Cell Cycle of Dictyostelium Discoideum in Liquid Nutrient MediumSandhya MadhavanPas encore d'évaluation
- Said AhmedDocument19 pagesSaid Ahmedahmed mahdiPas encore d'évaluation
- LactosacarosaDocument8 pagesLactosacarosaAriel plutarco cardoso herreraPas encore d'évaluation
- Role of Reactive Oxygen Species in p53 Activation During Cisplatin-Induced Apoptosis of Rat Mesangial CellsDocument7 pagesRole of Reactive Oxygen Species in p53 Activation During Cisplatin-Induced Apoptosis of Rat Mesangial CellsPandji 'Utuh'Pas encore d'évaluation
- Manoalide, Natural Sesterterpenoid That Inhibits Calcium Channels"Document8 pagesManoalide, Natural Sesterterpenoid That Inhibits Calcium Channels"Collin LoganPas encore d'évaluation
- Jsir 61 (9) 690-704 PDFDocument15 pagesJsir 61 (9) 690-704 PDFAusteridad LopezPas encore d'évaluation
- Vitamin C AutophagyDocument12 pagesVitamin C AutophagyureehwsenqPas encore d'évaluation
- LFABP and HFABPDocument6 pagesLFABP and HFABPMichiko MatsuoPas encore d'évaluation
- Use Only: New in Vitro Dna Polymerase Activity and Fidelity Assay Reveals Age-Dependent Changes in Arabidopsis ThalianaDocument6 pagesUse Only: New in Vitro Dna Polymerase Activity and Fidelity Assay Reveals Age-Dependent Changes in Arabidopsis ThalianacroitorugeorgianaPas encore d'évaluation
- 2011 Article 9671Document9 pages2011 Article 9671Brandon GriffinPas encore d'évaluation
- Wang 1996Document7 pagesWang 1996Anh NguyenPas encore d'évaluation
- Effect of Quercetin On Paraoxonase 1 Activity - Studies in Cultured Cells, Mice and HumansDocument7 pagesEffect of Quercetin On Paraoxonase 1 Activity - Studies in Cultured Cells, Mice and Humansanisa silviPas encore d'évaluation
- Ribosome-Inactivating Protein and Apoptosis: Abrin Causes Cell Death Via Mitochondrial Pathway in Jurkat CellsDocument8 pagesRibosome-Inactivating Protein and Apoptosis: Abrin Causes Cell Death Via Mitochondrial Pathway in Jurkat CellsKusuma WardaniPas encore d'évaluation
- NIH Public Access: Author ManuscriptDocument14 pagesNIH Public Access: Author ManuscriptYEME7HPas encore d'évaluation
- البحث السادسDocument11 pagesالبحث السادسsoha ahmedPas encore d'évaluation
- Cloning and Heterologous Expression of Hematin-Dependent Catalase Produced by Lactobacillus Plantarum CNRZ 1228Document4 pagesCloning and Heterologous Expression of Hematin-Dependent Catalase Produced by Lactobacillus Plantarum CNRZ 1228RickyAzrofiSamaraPas encore d'évaluation
- 1 s2.0 S0968089607009078 MainDocument10 pages1 s2.0 S0968089607009078 MainAirton Roman AguilarPas encore d'évaluation
- 02JaChroA OPANDADocument8 pages02JaChroA OPANDA이용문Pas encore d'évaluation
- Tian 2016Document13 pagesTian 2016Epiphany SonderPas encore d'évaluation
- Influence of Antidepressant Drugs On EctDocument7 pagesInfluence of Antidepressant Drugs On EctconstancePas encore d'évaluation
- KrsmanovickDocument7 pagesKrsmanovickKayro Lopez RamírezPas encore d'évaluation
- As 2. Genetics of Makapuno. A Genetic Tumor of The Coconut Endosperm Dolores A. Ramirez - (As)Document18 pagesAs 2. Genetics of Makapuno. A Genetic Tumor of The Coconut Endosperm Dolores A. Ramirez - (As)czelleah de graciaPas encore d'évaluation
- J Med Chem 2004 47 6609-6615-OKDocument7 pagesJ Med Chem 2004 47 6609-6615-OKca_teixeiraPas encore d'évaluation
- Doxi en Superficie OcularDocument7 pagesDoxi en Superficie OcularAna AloisiPas encore d'évaluation
- Insulin Secretion From Perifused Rat Pancreatic PseudoisletsDocument7 pagesInsulin Secretion From Perifused Rat Pancreatic PseudoisletskalwusaidiPas encore d'évaluation
- Isolation and Analysis of Nephritic Producing Immune Complexes in Plasmodium Berghei Infected MiceDocument7 pagesIsolation and Analysis of Nephritic Producing Immune Complexes in Plasmodium Berghei Infected MicedarknigterPas encore d'évaluation
- Journal of Neurochemistry Doi: 10.1111/j.1471-4159.2008.05762.xDocument13 pagesJournal of Neurochemistry Doi: 10.1111/j.1471-4159.2008.05762.xEdith ChaguaPas encore d'évaluation
- Liver Amylase: Iii. Synthesis by The Perfused Liver and Secretion Into The Perfusion MediumDocument6 pagesLiver Amylase: Iii. Synthesis by The Perfused Liver and Secretion Into The Perfusion MediumSantiago José pardoPas encore d'évaluation
- Shu - 2009 Aspergillus EstruturaDocument8 pagesShu - 2009 Aspergillus EstruturaTatiane RibeiroPas encore d'évaluation
- Bi 0274322Document10 pagesBi 0274322Carlos MeirellesPas encore d'évaluation
- Biochemical Factors Concerned in the Functional Activity of the Nervous System: First International Meeting of the International Society for Neurochemistry, Strasbourg, 1967D'EverandBiochemical Factors Concerned in the Functional Activity of the Nervous System: First International Meeting of the International Society for Neurochemistry, Strasbourg, 1967D. RichterPas encore d'évaluation
- Keragaan Pedagang Makanan Jajanan Olahan Di Kampus Universitas LampungDocument9 pagesKeragaan Pedagang Makanan Jajanan Olahan Di Kampus Universitas LampungFauziana NurhanisahPas encore d'évaluation
- 1689 12 PDFDocument8 pages1689 12 PDFFauziana NurhanisahPas encore d'évaluation
- (23279834 - HortScience) Anticancer Properties of Fruits and Vegetables PDFDocument3 pages(23279834 - HortScience) Anticancer Properties of Fruits and Vegetables PDFFauziana NurhanisahPas encore d'évaluation
- 1 s2.0 S0958166918300740 Main PDFDocument12 pages1 s2.0 S0958166918300740 Main PDFFauziana NurhanisahPas encore d'évaluation
- Biotechnology Advances: Carla Oliveira, Pedro M.R. Guimarães, Lucília DominguesDocument10 pagesBiotechnology Advances: Carla Oliveira, Pedro M.R. Guimarães, Lucília DominguesFauziana NurhanisahPas encore d'évaluation
- The Production of Chronic Renal Disease Rats by Single Intravenous Injection of Aminonucleoside OF Puromycin Effect of TinuousDocument11 pagesThe Production of Chronic Renal Disease Rats by Single Intravenous Injection of Aminonucleoside OF Puromycin Effect of TinuousFauziana NurhanisahPas encore d'évaluation
- Focal Single Intravenous Aminonucleoside: Following Dose PuromycinDocument7 pagesFocal Single Intravenous Aminonucleoside: Following Dose PuromycinFauziana NurhanisahPas encore d'évaluation
- 2-Hydroxyestradiol Attenuates Renal Disease in Chronic Puromycin Aminonucleoside NephropathyDocument11 pages2-Hydroxyestradiol Attenuates Renal Disease in Chronic Puromycin Aminonucleoside NephropathyFauziana NurhanisahPas encore d'évaluation
- Some Effects of Actinomycin D, Cycloheximide and Puromycin On Cell AdhesionDocument10 pagesSome Effects of Actinomycin D, Cycloheximide and Puromycin On Cell AdhesionFauziana NurhanisahPas encore d'évaluation
- Andreslacueva2008 2 PDFDocument7 pagesAndreslacueva2008 2 PDFFauziana NurhanisahPas encore d'évaluation
- Development and Physical Analysis of High Fiber Bread Incorporated With Cocoa (Theobroma Cacao SP.) Pod Husk PowderDocument5 pagesDevelopment and Physical Analysis of High Fiber Bread Incorporated With Cocoa (Theobroma Cacao SP.) Pod Husk PowderFauziana NurhanisahPas encore d'évaluation
- Usefulness of Thermo Labile Alkaline Phosphatase As A Biomarker in Diagnosis of Post-Menopausal OsteoporosisDocument6 pagesUsefulness of Thermo Labile Alkaline Phosphatase As A Biomarker in Diagnosis of Post-Menopausal OsteoporosisIJAR JOURNALPas encore d'évaluation
- Enzymes Biochemie VorlesungDocument48 pagesEnzymes Biochemie VorlesungbrokolliPas encore d'évaluation
- Liver Function Tests: Presented By-Dr. Arshiya Kaura (JR 1)Document50 pagesLiver Function Tests: Presented By-Dr. Arshiya Kaura (JR 1)vikasPas encore d'évaluation
- Year 2: PBL 2: JaundiceDocument74 pagesYear 2: PBL 2: JaundiceyaneemayPas encore d'évaluation
- Enzyme IsoenzymesDocument60 pagesEnzyme IsoenzymesSrishti GoenkaPas encore d'évaluation
- Validation of Milk Product Pasteurization by CDVS. 2018Document12 pagesValidation of Milk Product Pasteurization by CDVS. 2018Anonymous zIj5DCCAPas encore d'évaluation
- Mindray BA 88ADocument38 pagesMindray BA 88Aanisa nur insani100% (1)
- Spintrol "H" CAL: Human Serum / Suero HumanoDocument4 pagesSpintrol "H" CAL: Human Serum / Suero HumanoMD ALAMINPas encore d'évaluation
- Hepatoprotective Activity of Ocimum Americanum L LDocument7 pagesHepatoprotective Activity of Ocimum Americanum L LNurul Rachman NasutionPas encore d'évaluation
- (Hong Et Al 2012) Effects of Fermented Black Soybean Pulp On Lipid and Bone Metabolism in Ovariectomized RatsDocument7 pages(Hong Et Al 2012) Effects of Fermented Black Soybean Pulp On Lipid and Bone Metabolism in Ovariectomized RatsSie ningsihPas encore d'évaluation
- Nigella Sativa (Black Seed) Safety: An Overview: Open Access ReviewDocument11 pagesNigella Sativa (Black Seed) Safety: An Overview: Open Access ReviewShinta DamayantiPas encore d'évaluation
- CC 2 LAB - ALP and ACPDocument7 pagesCC 2 LAB - ALP and ACPAngelaAmmcoPas encore d'évaluation
- Multidimensional Uses of Medicinal Plantkachnar Bauhinia Variegata LinnDocument15 pagesMultidimensional Uses of Medicinal Plantkachnar Bauhinia Variegata Linnkeshav shishyaPas encore d'évaluation
- Enzymes: M. Zaharna Clin. Chem. 2009Document32 pagesEnzymes: M. Zaharna Clin. Chem. 2009Ahmed GaberPas encore d'évaluation
- Liver Function Tests - Purpose and ProcedureDocument12 pagesLiver Function Tests - Purpose and ProcedureJennifer CollinsPas encore d'évaluation
- Standard LFT and It's Clinical SignificanceDocument28 pagesStandard LFT and It's Clinical SignificanceanimathzPas encore d'évaluation
- c311 ALP2 enDocument3 pagesc311 ALP2 endr. SheryarOrakzaiPas encore d'évaluation
- Measurement of EnzymesDocument71 pagesMeasurement of EnzymesCJ VillanuevaPas encore d'évaluation
- Clinical Chemistry 2Document22 pagesClinical Chemistry 2Rubenne Miles ElagasPas encore d'évaluation
- Alkaline Phosphatase - Dimension - Rev C DXDCM 09017fe980338e46-1559358386133Document11 pagesAlkaline Phosphatase - Dimension - Rev C DXDCM 09017fe980338e46-1559358386133Paolo Bolvaran JulioPas encore d'évaluation
- Clinically Important EnzymesDocument29 pagesClinically Important Enzymesonnumilla49Pas encore d'évaluation
- Procedure Catalogue PhoenixDocument203 pagesProcedure Catalogue PhoenixVinsmoke SanjiPas encore d'évaluation
- Safety Evaluation of Mercury Based Ayurvedic Formulation (Sidh Makardhwaj) On Brain Cerebrum, Liver & Kidney in RatsDocument10 pagesSafety Evaluation of Mercury Based Ayurvedic Formulation (Sidh Makardhwaj) On Brain Cerebrum, Liver & Kidney in RatsSaliniPas encore d'évaluation
- ALP2 0003333752190COIN V5 enDocument4 pagesALP2 0003333752190COIN V5 enMMCSTOREPas encore d'évaluation
- 179-Article Text-302-1-10-20171121 PDFDocument5 pages179-Article Text-302-1-10-20171121 PDFMichael XuPas encore d'évaluation
- Alkaline Phosphatase: 4 X 90/4 X 26 ML 12011003Document1 pageAlkaline Phosphatase: 4 X 90/4 X 26 ML 12011003Aniket DubeyPas encore d'évaluation
- Alkaline Phosphatase: Test Principle: Enzymatic ColorimetricDocument2 pagesAlkaline Phosphatase: Test Principle: Enzymatic ColorimetricValdez Francis ZaccheauPas encore d'évaluation
- Morphological, Haematological and Biochemical Changes in African Catfish Clarias Gariepinus (Burchell 1822) Juveniles Exposed To ClotrimazoleDocument7 pagesMorphological, Haematological and Biochemical Changes in African Catfish Clarias Gariepinus (Burchell 1822) Juveniles Exposed To ClotrimazoleAndres Fernando Silvestre SuarezPas encore d'évaluation
- Protocolos DiaSys Stardust MC15Document61 pagesProtocolos DiaSys Stardust MC15leopa78100% (1)
- Theories of MineralizationDocument1 pageTheories of Mineralizationanku_2426Pas encore d'évaluation