Vous aimerez peut-être aussi
- Guia Aceites EsencialesDocument72 pagesGuia Aceites Esencialesrubis Carlos Araujo91% (11)
- Anatomía FaringeDocument27 pagesAnatomía FaringeGary David Morales Rodriguez100% (1)
- Derecho Penal Parte Especial-Tomo I-Tomas Galvez Villegas y Ricardo Rojas LeonDocument1 088 pagesDerecho Penal Parte Especial-Tomo I-Tomas Galvez Villegas y Ricardo Rojas Leonnomejoas enserio67% (3)
- Apuntes de Materia y EnergiaDocument42 pagesApuntes de Materia y EnergiaSaul CalderonPas encore d'évaluation
- DIENCEFALO GaryDocument46 pagesDIENCEFALO GaryGary David Morales Rodriguez100% (1)
- 1 Nivel de Org CorpDocument9 pages1 Nivel de Org CorpGary David Morales RodriguezPas encore d'évaluation
- Nervios CranealesDocument50 pagesNervios CranealesGary David Morales RodriguezPas encore d'évaluation
- ToraxDocument35 pagesToraxGary David Morales RodriguezPas encore d'évaluation
- Pares CranealesDocument12 pagesPares CranealesGary David Morales RodriguezPas encore d'évaluation
- Anatomía de La BocaDocument24 pagesAnatomía de La BocaGary David Morales RodriguezPas encore d'évaluation
- Malformaciones Del Aparato RespiratorioDocument22 pagesMalformaciones Del Aparato RespiratorioGary David Morales RodriguezPas encore d'évaluation
- Vías de AdministraciónDocument3 pagesVías de AdministraciónGary David Morales RodriguezPas encore d'évaluation
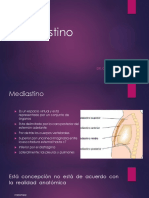
- MediastinoDocument11 pagesMediastinoGary David Morales RodriguezPas encore d'évaluation
- Anatomía Aparato Genital MasculinoDocument37 pagesAnatomía Aparato Genital MasculinoGary David Morales RodriguezPas encore d'évaluation
- Control y Evaluación Del EscenarioDocument4 pagesControl y Evaluación Del EscenarioGary David Morales RodriguezPas encore d'évaluation
- Musculos Miembro SuperiorDocument48 pagesMusculos Miembro SuperiorGary David Morales RodriguezPas encore d'évaluation
- Glosofaríngeo IX Par CranealDocument12 pagesGlosofaríngeo IX Par CranealGary David Morales RodriguezPas encore d'évaluation
- Vias de AdmiDocument27 pagesVias de AdmiGary David Morales RodriguezPas encore d'évaluation
- Guia de Administracion de MedicamentosDocument2 pagesGuia de Administracion de MedicamentosGary David Morales RodriguezPas encore d'évaluation
- GL OsarioDocument4 pagesGL OsarioGary David Morales RodriguezPas encore d'évaluation
- Terminología MédicaDocument19 pagesTerminología MédicaGary David Morales RodriguezPas encore d'évaluation
- Terminología MédicaDocument19 pagesTerminología MédicaGary David Morales RodriguezPas encore d'évaluation
- Manual VEDocument40 pagesManual VEGary David Morales RodriguezPas encore d'évaluation
- Traumatismos en ExtremidadesDocument4 pagesTraumatismos en ExtremidadesGary David Morales RodriguezPas encore d'évaluation
- Heridas y Control de HemorragiasDocument5 pagesHeridas y Control de HemorragiasGary David Morales RodriguezPas encore d'évaluation
- Encuesta EpidemiologicaDocument4 pagesEncuesta EpidemiologicaGary David Morales Rodriguez0% (1)
- Obstrucción de Vía AéreaDocument27 pagesObstrucción de Vía AéreaGary David Morales RodriguezPas encore d'évaluation
- Critica A La EducacionDocument2 pagesCritica A La EducacionGary David Morales RodriguezPas encore d'évaluation
- Tarea Brotes GaryDocument22 pagesTarea Brotes GaryGary David Morales RodriguezPas encore d'évaluation
- Heridas y Control de HemorragiasDocument5 pagesHeridas y Control de HemorragiasGary David Morales RodriguezPas encore d'évaluation
- Primera SemanaDocument24 pagesPrimera SemanaGary David Morales RodriguezPas encore d'évaluation
- Tercer Mes Hasta La Fecha Del PartoDocument10 pagesTercer Mes Hasta La Fecha Del PartoGary David Morales RodriguezPas encore d'évaluation
- Apuntes de EnfermeriaDocument8 pagesApuntes de EnfermeriaJoana TorresPas encore d'évaluation
- Mantenimiento 55000Document2 pagesMantenimiento 55000Christian Ruben Valera VillenaPas encore d'évaluation
- Rastros Municipales Final 04Document80 pagesRastros Municipales Final 04Daniel Cruz100% (1)
- Sara Maria AldreteDocument4 pagesSara Maria AldretecfpsaroPas encore d'évaluation
- Alfa Laval SPC 2 Valvula Reguladora Folletos Del ProductoDocument4 pagesAlfa Laval SPC 2 Valvula Reguladora Folletos Del ProductoJose GalindoPas encore d'évaluation
- Mejora de Servicio BrinsaDocument19 pagesMejora de Servicio Brinsafredy ramirezPas encore d'évaluation
- 4 Lumholtz Arte SimbólicoDocument6 pages4 Lumholtz Arte SimbólicokDs KaraPas encore d'évaluation
- Protocolo Regularización Del RebordeDocument6 pagesProtocolo Regularización Del RebordeFernanda MadrigalPas encore d'évaluation
- SIMPOSIO 1 Psico-neuro-pedagogía-Miguel Zubiria 1Document27 pagesSIMPOSIO 1 Psico-neuro-pedagogía-Miguel Zubiria 1Dorian Miguel Ospino CaroPas encore d'évaluation
- Tratamiento Del AsmaDocument8 pagesTratamiento Del AsmaDenis José FernándezPas encore d'évaluation
- Guia Rapida VAT - V2Document13 pagesGuia Rapida VAT - V2Marco Viza100% (1)
- Raviolis de Gambas Con NageDocument4 pagesRaviolis de Gambas Con Nagemarcos gonzalezPas encore d'évaluation
- Habitos y Consumo de Las Bebidas RehidratantesDocument31 pagesHabitos y Consumo de Las Bebidas Rehidratanteshugo chungaPas encore d'évaluation
- Proyecto Bachillerato JosefinoDocument17 pagesProyecto Bachillerato JosefinoEdgar RangelPas encore d'évaluation
- Potencial Receptor - Grupo #4Document16 pagesPotencial Receptor - Grupo #4Albert Christopher Vargas QuirozPas encore d'évaluation
- Perfil Del Proyecto MINA POOPO Min-3911 Vargas Hurtado Jorge LuisDocument39 pagesPerfil Del Proyecto MINA POOPO Min-3911 Vargas Hurtado Jorge LuisDaniela PortilloPas encore d'évaluation
- Relectura BíblicaDocument23 pagesRelectura BíblicarlarroyavecPas encore d'évaluation
- Anexo 1 Ficha para Análisis de Lectura (FINAL)Document19 pagesAnexo 1 Ficha para Análisis de Lectura (FINAL)Karol JaramilloPas encore d'évaluation
- Taller #1 LácteosDocument3 pagesTaller #1 LácteosNataly NuñezPas encore d'évaluation
- Practica N°3Document11 pagesPractica N°3FudoPas encore d'évaluation
- Cuestionario - Mentes Diferentes, Aprendizajes DiferentesDocument3 pagesCuestionario - Mentes Diferentes, Aprendizajes DiferentesA. H. Toledo, L.Psic.Pas encore d'évaluation
- Auditoria Evidencias y HallazgosDocument12 pagesAuditoria Evidencias y HallazgosCristopher MartinezPas encore d'évaluation
- Visita Al Molino AmazonasDocument11 pagesVisita Al Molino Amazonasanon_2482762010% (1)
- OriginalyDocument40 pagesOriginalyjohana9142Pas encore d'évaluation
- Proyecto Vial (Jorge Darren Piscoche Cenizario)Document24 pagesProyecto Vial (Jorge Darren Piscoche Cenizario)Jorge D. Piscoche CenizarioPas encore d'évaluation
- 01.07 Red Distribucion .ActualizadoDocument3 pages01.07 Red Distribucion .Actualizadopedro sucasacaPas encore d'évaluation
- Curación de Heridas QuirurgicasDocument28 pagesCuración de Heridas QuirurgicasGemaElizabethSaltosPas encore d'évaluation