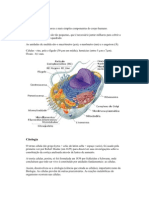
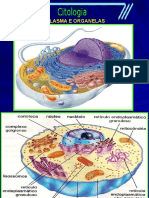
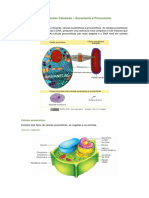
Vous aimerez peut-être aussi
- Aula 5Document13 pagesAula 5Aniceto RosarioPas encore d'évaluation
- AULA Organelas CitoplasmáticasDocument20 pagesAULA Organelas CitoplasmáticasJoana TalitaPas encore d'évaluation
- Introdução A BioquímicaDocument6 pagesIntrodução A BioquímicaDeborah RochaPas encore d'évaluation
- CITOLOGIA Classificação Das CelulasDocument53 pagesCITOLOGIA Classificação Das Celulascamila.alvesPas encore d'évaluation
- Citologia Revisado Pontos Importantes Aula de BiologiaDocument2 pagesCitologia Revisado Pontos Importantes Aula de BiologiaYara FerroPas encore d'évaluation
- Livro Fisiologia Básica PDFDocument8 pagesLivro Fisiologia Básica PDFSimone KrupokPas encore d'évaluation
- ICM - Biologia - Parte 2Document55 pagesICM - Biologia - Parte 2Edmond TPas encore d'évaluation
- Citoplasma e Organelas CitoplasmáticasDocument46 pagesCitoplasma e Organelas Citoplasmáticas0774.rosimeribatistaPas encore d'évaluation
- 205949-Aula 2 OrganelasDocument22 pages205949-Aula 2 Organelaskeyse rochaPas encore d'évaluation
- Citoplasma, HialoplasmaDocument40 pagesCitoplasma, HialoplasmaJordi GeraldesPas encore d'évaluation
- Retículo EndoplasmáticoDocument10 pagesRetículo EndoplasmáticoWillyane Samara Leandro FerreiraPas encore d'évaluation
- Aula 05 Sistemas de Endomembranas e OrganelasDocument24 pagesAula 05 Sistemas de Endomembranas e OrganelasBIANCA PIFFERPas encore d'évaluation
- Resumo - GenéticaDocument16 pagesResumo - Genéticaisabeljulia968Pas encore d'évaluation
- A Base Cromossômica Aula 1Document23 pagesA Base Cromossômica Aula 1Maria Clara FernandesPas encore d'évaluation
- Imprimir - Organelas CitoplasmaticasDocument29 pagesImprimir - Organelas Citoplasmaticassantosalanne5Pas encore d'évaluation
- Biologia Aula 2Document8 pagesBiologia Aula 2Kelly DomboPas encore d'évaluation
- Biologia Aula 2Document8 pagesBiologia Aula 2Pierry IsseiPas encore d'évaluation
- CelulaDocument28 pagesCelulaJorge William NhampossePas encore d'évaluation
- Apostila Citologia e HistologiaDocument23 pagesApostila Citologia e HistologiaFlor DoSolPas encore d'évaluation
- INTRODUÇÃO A FISIOLOGIA (Biologia Celular)Document19 pagesINTRODUÇÃO A FISIOLOGIA (Biologia Celular)geojac2112Pas encore d'évaluation
- Aula 3 CitologiaDocument42 pagesAula 3 CitologiaPapelaria TeixeiraPas encore d'évaluation
- Resumo Organelas CitoplasmáticasDocument2 pagesResumo Organelas CitoplasmáticasCaroline Angeli Sancio100% (1)
- Célula AnimalDocument6 pagesCélula AnimalFelipe MeloPas encore d'évaluation
- Biologia CelularDocument9 pagesBiologia CelularLeonardo MorgadoPas encore d'évaluation
- Aula 1.1 - Citoplasma e Organelas CitoplasmáticasDocument35 pagesAula 1.1 - Citoplasma e Organelas CitoplasmáticasFeel Oliver100% (1)
- Aula 7 - Organelas Citoplasmáticas IIDocument37 pagesAula 7 - Organelas Citoplasmáticas IIhome maiaPas encore d'évaluation
- Aula 5 - Citoplasma e OrganelasDocument24 pagesAula 5 - Citoplasma e OrganelasUrias NobrePas encore d'évaluation
- A Célula PDFDocument8 pagesA Célula PDFGildaMarisaPas encore d'évaluation
- Atividade Contextualizada Celula Eucarionte e FunçõesDocument5 pagesAtividade Contextualizada Celula Eucarionte e FunçõesjosewillyPas encore d'évaluation
- Membrana BiologicaDocument5 pagesMembrana BiologicaMaria Eduarda Padilha BaltazarPas encore d'évaluation
- Organelas CelularesDocument6 pagesOrganelas CelularesLuara Patrine SalesPas encore d'évaluation
- Aula 2 CitoplasmaDocument38 pagesAula 2 CitoplasmaJoice Caroline Lucas JuniorPas encore d'évaluation
- Caderno - Bases BiológicasDocument3 pagesCaderno - Bases BiológicasFabiano ViannaPas encore d'évaluation
- CITOPLASMADocument89 pagesCITOPLASMAVagner Fortes RosadoPas encore d'évaluation
- Organitos e Suas Funções - JR - para PreencherDocument2 pagesOrganitos e Suas Funções - JR - para PreencherMariana AnjosPas encore d'évaluation
- Objetivos Respondidos (Biomoléculas I, II, III, IV e V, e MeDocument24 pagesObjetivos Respondidos (Biomoléculas I, II, III, IV e V, e MeWinnie ChitundaPas encore d'évaluation
- Citoplasma e OrganelasDocument23 pagesCitoplasma e OrganelasMariaNavesRibeiroLopesPas encore d'évaluation
- Ridiculo Endoplasmatico FalasDocument4 pagesRidiculo Endoplasmatico FalasMadalena SantosPas encore d'évaluation
- Organelas Citoplasmáticas-1Document12 pagesOrganelas Citoplasmáticas-1Mary ReisPas encore d'évaluation
- Citologia BásicaDocument6 pagesCitologia BásicaAna Clara SilvaPas encore d'évaluation
- Citologia 12classe 22Document11 pagesCitologia 12classe 22Beto AgostinhoPas encore d'évaluation
- Av1 - BiologiaDocument7 pagesAv1 - BiologiaLea SousaPas encore d'évaluation
- Organelas CelulareDocument11 pagesOrganelas CelulareGabriel PrataPas encore d'évaluation
- CitologiaDocument7 pagesCitologiaTeresa Rosaria de Souza Gonzalez EnriquezPas encore d'évaluation
- Trabalho de BCM FinalDocument19 pagesTrabalho de BCM FinalKieza JoaquimPas encore d'évaluation
- Trabalho Biologia AmanhãDocument10 pagesTrabalho Biologia AmanhãVictor GiembraPas encore d'évaluation
- Biologia PPT - Citoplasma - OrganelasDocument13 pagesBiologia PPT - Citoplasma - OrganelasBiologia ppt100% (5)
- A CélulaDocument6 pagesA CélulaPedro GouveiaPas encore d'évaluation
- 1celula NormalDocument66 pages1celula NormalSamuel Gomes SouzaPas encore d'évaluation
- Aula 3 Citoplasma e OrganelasDocument37 pagesAula 3 Citoplasma e OrganelassilrogerioPas encore d'évaluation
- PESQUISADocument6 pagesPESQUISAsamara vitoria frias zanellato zanellatoPas encore d'évaluation
- Trabalho de Biologia CelularDocument6 pagesTrabalho de Biologia CelularLuiz Felipe SalomãoPas encore d'évaluation
- CitologiaDocument8 pagesCitologiaDenise CorreiaPas encore d'évaluation
- Estrutura Celular ProntoDocument4 pagesEstrutura Celular ProntoCauet CamargoPas encore d'évaluation
- Citoplasma e NúcleoDocument11 pagesCitoplasma e NúcleoAna Paula Porto FerreiraPas encore d'évaluation
- Resumo OrganelasDocument4 pagesResumo OrganelasLEONARDO MACIEL LIMAPas encore d'évaluation
- U3 - S1 - PÓS - Bio1Document4 pagesU3 - S1 - PÓS - Bio1QuemPas encore d'évaluation
- Resumos de Biologia e GeologiaDocument34 pagesResumos de Biologia e GeologiaMariana MeninoPas encore d'évaluation
- Estudo Dirigido Sinalização e Diferenciação Celular PDFDocument1 pageEstudo Dirigido Sinalização e Diferenciação Celular PDFAvguzto Plinio Souza100% (1)
- Síntese de Ácidos GraxosDocument9 pagesSíntese de Ácidos Graxosfernandoribeirojr98Pas encore d'évaluation
- Caderno de Questões Introdução A FarmacologiaDocument16 pagesCaderno de Questões Introdução A FarmacologiaJair MoreiraPas encore d'évaluation
- Resumo Neurociências SensorialDocument13 pagesResumo Neurociências SensorialRianne GomesPas encore d'évaluation
- Caderno de Questões 1Document44 pagesCaderno de Questões 1Adriel LimaPas encore d'évaluation
- UFAC 2014dDocument16 pagesUFAC 2014dLUCES Liga Universitaria de Cirugías y EstéticaPas encore d'évaluation
- Patologias Associadas As Vias de Transdução de SinalDocument22 pagesPatologias Associadas As Vias de Transdução de SinalMilla LuckPas encore d'évaluation
- Fisiologia Endócrina E Do Metabolismo: Prof Beatriz Essenfelder BorgesDocument96 pagesFisiologia Endócrina E Do Metabolismo: Prof Beatriz Essenfelder Borgeslii pereiraPas encore d'évaluation
- FARMACOLOGIADocument84 pagesFARMACOLOGIALarissa Ayashi KawanichiPas encore d'évaluation
- Aula Interação Fármaco-Receptor PDFDocument61 pagesAula Interação Fármaco-Receptor PDFLuciara RochaPas encore d'évaluation
- 2 Aula FarmadinamicaDocument48 pages2 Aula FarmadinamicaEduardo CostaPas encore d'évaluation
- Mecanismos de Transdução de Sinal para Os Receptores de Membrana PlasmáticaDocument12 pagesMecanismos de Transdução de Sinal para Os Receptores de Membrana PlasmáticaJacqueline Quintal100% (1)
- Transtorno BipolarDocument30 pagesTranstorno BipolardesireepaysagePas encore d'évaluation
- 029 Biossinalizacao 009Document41 pages029 Biossinalizacao 009Manuela M. Lins100% (1)
- Receptores Farmacológicos e Sistemas EfetoresDocument9 pagesReceptores Farmacológicos e Sistemas EfetoresEmilly EduardaPas encore d'évaluation
- Olfação e Gustação 28 Abr 2023 Alex Kors Vidsiunas-1Document18 pagesOlfação e Gustação 28 Abr 2023 Alex Kors Vidsiunas-1minato.namikaze2901Pas encore d'évaluation
- Aula 1Document44 pagesAula 1Marina SalesPas encore d'évaluation
- Farmacologia BiomedicinaDocument5 pagesFarmacologia Biomedicinaedusoares30Pas encore d'évaluation
- Farmacocinetica e FarmacodinamicaDocument19 pagesFarmacocinetica e FarmacodinamicaPablo AlbinoPas encore d'évaluation
- Vias de Administração e FarmacodinamicaDocument26 pagesVias de Administração e FarmacodinamicaZoePas encore d'évaluation
- Circulação de Leucócitos e Migração para Os TecidosDocument18 pagesCirculação de Leucócitos e Migração para Os TecidosVitoria Pires FelixPas encore d'évaluation
- Guia de Referência Do Sistema Nervoso - DIGITALDocument136 pagesGuia de Referência Do Sistema Nervoso - DIGITALnaemi ishiguroPas encore d'évaluation
- Mecanismos de Comunicação CelularDocument10 pagesMecanismos de Comunicação CelularBruno CordeiroPas encore d'évaluation
- Material Estudo Transdução SensorialDocument13 pagesMaterial Estudo Transdução SensorialMichele Perrone FilhoPas encore d'évaluation
- Sinalização CelularDocument41 pagesSinalização CelularAna BarthPas encore d'évaluation
- Apostila Aulas Praticas Farmaco UnifespDocument26 pagesApostila Aulas Praticas Farmaco UnifespmariliapatraoPas encore d'évaluation
- FARMACOLOGIADocument14 pagesFARMACOLOGIAAlexandre Assis do CarmoPas encore d'évaluation
- Farmacologia Resumo PDFDocument14 pagesFarmacologia Resumo PDFbeaPas encore d'évaluation
- BiofísicaDocument4 pagesBiofísicaVictoria MendesPas encore d'évaluation
- Farmacodinâmica PDFDocument45 pagesFarmacodinâmica PDFClovis CarvalhoPas encore d'évaluation