Vous aimerez peut-être aussi
- Cysteine Cathepsins in Human Dentin-Pulp ComplexDocument7 pagesCysteine Cathepsins in Human Dentin-Pulp ComplexAfonso OtsukaPas encore d'évaluation
- Amelogenins: Multi-Functional Proteins in Enamel & Periodontal TissuesDocument13 pagesAmelogenins: Multi-Functional Proteins in Enamel & Periodontal TissuesSABUESO FINANCIEROPas encore d'évaluation
- Matrix Metalloproteinase-8 (MMP-8) Is The Major Collagenase in Human DentinDocument7 pagesMatrix Metalloproteinase-8 (MMP-8) Is The Major Collagenase in Human DentinedelinPas encore d'évaluation
- Alteration of Enamel Proteins in Hypomaturation Amelogenesis ImperfectaDocument3 pagesAlteration of Enamel Proteins in Hypomaturation Amelogenesis ImperfectaVerónica RuizPas encore d'évaluation
- Micro-Dissection of Enamel Organ From Mandibular Incisor of Rats Exposed To Environmental ToxicantsDocument10 pagesMicro-Dissection of Enamel Organ From Mandibular Incisor of Rats Exposed To Environmental ToxicantsNadira NurinPas encore d'évaluation
- Mu 2017Document14 pagesMu 2017Jorge Esteban Briceño MoyaPas encore d'évaluation
- The Dental Pulp: Biology, Pathology, and Regenerative TherapiesD'EverandThe Dental Pulp: Biology, Pathology, and Regenerative TherapiesPas encore d'évaluation
- Amelogenin Phosphorylation Regulates Tooth Enamel Formation by Stabilizing A Transient Amorphous Mineral PrecursorDocument17 pagesAmelogenin Phosphorylation Regulates Tooth Enamel Formation by Stabilizing A Transient Amorphous Mineral PrecursorratrikaruniaPas encore d'évaluation
- The Effects of Mineral Trioxide Aggregates On Cytokine Production by Mouse Pulp TissueDocument7 pagesThe Effects of Mineral Trioxide Aggregates On Cytokine Production by Mouse Pulp TissueOmar DiazPas encore d'évaluation
- Ameloblastin Is Not Implicated in Bone Remodelling and RepairDocument13 pagesAmeloblastin Is Not Implicated in Bone Remodelling and RepairZi JianPas encore d'évaluation
- Mast Cells and MMP 9 in The Lamina ProprDocument10 pagesMast Cells and MMP 9 in The Lamina ProprLuciana CarvalhoPas encore d'évaluation
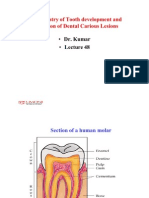
- Biochemistry of tooth development and formation of dental carious lesionsDocument31 pagesBiochemistry of tooth development and formation of dental carious lesionsosamamusaPas encore d'évaluation
- Effect of Antibiotics and NSAIDs On Cyclooxygenase-2 in The Enamel MineralizationDocument7 pagesEffect of Antibiotics and NSAIDs On Cyclooxygenase-2 in The Enamel MineralizationLjubomirErdoglijaPas encore d'évaluation
- 1 Repiska 2010Document6 pages1 Repiska 2010Thamyres BrancoPas encore d'évaluation
- Primary Teeth ResoptionDocument9 pagesPrimary Teeth ResoptionAdi AdiPas encore d'évaluation
- Trimethyltin Chloride Induces Reactive Oxygen Species-Mediated ApoptosisDocument9 pagesTrimethyltin Chloride Induces Reactive Oxygen Species-Mediated ApoptosissalmaPas encore d'évaluation
- Ingeniería TisularDocument7 pagesIngeniería TisularLAURA DIAZ PALENCIAPas encore d'évaluation
- NIH Public Access: Author ManuscriptDocument12 pagesNIH Public Access: Author ManuscriptMaria-Lavinia HoinaruPas encore d'évaluation
- Associations between oral bacteria and caries in Kenyan teensDocument7 pagesAssociations between oral bacteria and caries in Kenyan teensgabyPas encore d'évaluation
- Life Sciences: A B A A A ADocument10 pagesLife Sciences: A B A A A AsovalaxPas encore d'évaluation
- Chlorhexidine Increases The Longevity of in Vivo Resin-Dentin BondsDocument6 pagesChlorhexidine Increases The Longevity of in Vivo Resin-Dentin BondsAli Al-QaysiPas encore d'évaluation
- Enamel Defects Reflect Perinatal Exposure To Bisphenol A Jedeon Et Al AmJP 2013Document11 pagesEnamel Defects Reflect Perinatal Exposure To Bisphenol A Jedeon Et Al AmJP 2013Danny LorakPas encore d'évaluation
- Art Biologia Oral ApoptosisDocument6 pagesArt Biologia Oral ApoptosisDaniela CastilloPas encore d'évaluation
- Tooth Wear As A Means To Quantify Intra-Specific Variations in Diet and Chewing Movements - Calandra Et Al 2016Document10 pagesTooth Wear As A Means To Quantify Intra-Specific Variations in Diet and Chewing Movements - Calandra Et Al 2016Bogdan HaiducPas encore d'évaluation
- Functional Proteomics To Exploit Genome Sequences: A. Donny StrosbergDocument6 pagesFunctional Proteomics To Exploit Genome Sequences: A. Donny StrosbergscribPas encore d'évaluation
- Activation of The AktmTOR Pathway in Dentigerous Cysts, Odontogenic Keratocysts, and AmeloblastomasDocument7 pagesActivation of The AktmTOR Pathway in Dentigerous Cysts, Odontogenic Keratocysts, and AmeloblastomasCamila CastiblancoPas encore d'évaluation
- Ultrastructure of Dental CalculusDocument7 pagesUltrastructure of Dental CalculusAndrea UrgilesPas encore d'évaluation
- 3.podoplanin Antibody MainDocument6 pages3.podoplanin Antibody MainNitin KhanduriPas encore d'évaluation
- Articolo 3Document16 pagesArticolo 3Marco BulseiPas encore d'évaluation
- Histopathological Variations in Ameloblastoma - Cases in A Tertiary Care Center of NepalDocument7 pagesHistopathological Variations in Ameloblastoma - Cases in A Tertiary Care Center of NepalDinesh YadavPas encore d'évaluation
- Novel Transcription Factor Correlated with Glomerular Proliferation and SclerosisDocument8 pagesNovel Transcription Factor Correlated with Glomerular Proliferation and SclerosisSarly FebrianaPas encore d'évaluation
- Calpain May Contribute To Hereditary Cataract Formation in SheepDocument7 pagesCalpain May Contribute To Hereditary Cataract Formation in SheepNovel RamadhaniPas encore d'évaluation
- Archives of Oral BiologyDocument5 pagesArchives of Oral BiologyZana Salsabila ChobitaPas encore d'évaluation
- Lactose Persistence Nihms778525Document26 pagesLactose Persistence Nihms778525kaleb16_2Pas encore d'évaluation
- AMBN2Document18 pagesAMBN2Lisa PrihastariPas encore d'évaluation
- Dietary On Dental Calculus 2018Document10 pagesDietary On Dental Calculus 2018Ronaldo PutraPas encore d'évaluation
- 2011 EffectsofHEMA TEGDMA PulpstemcellsDocument11 pages2011 EffectsofHEMA TEGDMA Pulpstemcells팜브엉닥 / 학생 / 화학생물공학부Pas encore d'évaluation
- Recuperarea Defectului de Dimensiune Critică Al Osului Parietal La Iepure Prin Grefe CompoziteDocument7 pagesRecuperarea Defectului de Dimensiune Critică Al Osului Parietal La Iepure Prin Grefe CompoziteRoxana VoicuPas encore d'évaluation
- Cysteine Cathepsins in HumanDocument6 pagesCysteine Cathepsins in HumanAfonso OtsukaPas encore d'évaluation
- Uncaria Tomentosa Reduces Osteoclastic Bone Loss in Vivo: PhytomedicineDocument9 pagesUncaria Tomentosa Reduces Osteoclastic Bone Loss in Vivo: PhytomedicineGallardo JahsePas encore d'évaluation
- 1 s2.0 S0143400411005339 Main PDFDocument8 pages1 s2.0 S0143400411005339 Main PDFMarco Antonio AspronPas encore d'évaluation
- 2022 Papers Merged 15 23Document9 pages2022 Papers Merged 15 23Yeannie YapPas encore d'évaluation
- Odontoblast-Like Cell Differentiation and Dentin Formation Induced With TGF-b1Document9 pagesOdontoblast-Like Cell Differentiation and Dentin Formation Induced With TGF-b1juanesteban1890Pas encore d'évaluation
- Ingram JME09 LCT EthiopiaDocument11 pagesIngram JME09 LCT EthiopiaJosue BarralPas encore d'évaluation
- 1 s2.0 S0099239909002386 MainDocument5 pages1 s2.0 S0099239909002386 MainMunish BatraPas encore d'évaluation
- Biochimie: Angela C. Mora Huertas, Christian E.H. Schmelzer, Wolfgang Hoehenwarter, Frank Heyroth, Andrea HeinzDocument11 pagesBiochimie: Angela C. Mora Huertas, Christian E.H. Schmelzer, Wolfgang Hoehenwarter, Frank Heyroth, Andrea HeinzUtari DewiPas encore d'évaluation
- 6 Alaluusua2010Document6 pages6 Alaluusua2010Nicole StoicaPas encore d'évaluation
- 4NQO-Induced Rat Tongue Carcinoma: An Ultrastructural StudyDocument8 pages4NQO-Induced Rat Tongue Carcinoma: An Ultrastructural StudyGlauce L TrevisanPas encore d'évaluation
- J Dent Res 2011 Takahashi 1463 8Document6 pagesJ Dent Res 2011 Takahashi 1463 8abcder1234Pas encore d'évaluation
- Gelatinase A (MMP-2) in Developing Tooth Tissues and Amelogenin HydrolysisDocument5 pagesGelatinase A (MMP-2) in Developing Tooth Tissues and Amelogenin HydrolysisKairós OEPas encore d'évaluation
- 32 ND JC - SindhuDocument42 pages32 ND JC - SindhuDadi SindhuPas encore d'évaluation
- ArticuloDocument7 pagesArticuloCarlos LicéagaPas encore d'évaluation
- Celulas Madre en Los Dientes-Dental Cell Type Atlas Reveals Stem and Differentiated Cell Types in Mouse and Human TeethDocument18 pagesCelulas Madre en Los Dientes-Dental Cell Type Atlas Reveals Stem and Differentiated Cell Types in Mouse and Human TeethSusana FariasPas encore d'évaluation
- Deciphering Stem Cell From Apical Papilla– MDocument17 pagesDeciphering Stem Cell From Apical Papilla– Matif nawazPas encore d'évaluation
- Identification of Live Germ-Cell Desquamation As A Major Mechanism of SeasonalDocument12 pagesIdentification of Live Germ-Cell Desquamation As A Major Mechanism of SeasonalEstrella BenitoPas encore d'évaluation
- ODONTOGENESISDocument562 pagesODONTOGENESISanonimoPas encore d'évaluation
- Role of adipokines in cranial developmentDocument3 pagesRole of adipokines in cranial developmentzeljkostojakovicPas encore d'évaluation
- 2022 Dentaldefects Environmental EHP10208Document12 pages2022 Dentaldefects Environmental EHP10208Senussi IbtisamPas encore d'évaluation
- Inhibition of Orthodontically Induced Root Resorption With Echistatin, An RGD-containing PeptideDocument9 pagesInhibition of Orthodontically Induced Root Resorption With Echistatin, An RGD-containing PeptideAndres CoboPas encore d'évaluation
- Biocompatibility of Three Calcium Silicate Based Materials Implanted in Ratsubcutaneous TissueDocument9 pagesBiocompatibility of Three Calcium Silicate Based Materials Implanted in Ratsubcutaneous TissueRista LewiyonahPas encore d'évaluation
- Toxicologiacal Fluorides PDFDocument404 pagesToxicologiacal Fluorides PDFMoisés MotaPas encore d'évaluation
- A Jewish State - An Attempt at A Modern Solution of The Jewish QuestionDocument67 pagesA Jewish State - An Attempt at A Modern Solution of The Jewish QuestionbuhaptserenPas encore d'évaluation
- Secrets of LuciferDocument80 pagesSecrets of LuciferPedroMousinho96% (25)
- Exhibition of WATER COLORS by T. HERZL ROMEDocument4 pagesExhibition of WATER COLORS by T. HERZL ROMEMoisés MotaPas encore d'évaluation
- Brazil Trade Liberalization ProgramDocument18 pagesBrazil Trade Liberalization ProgramMoisés MotaPas encore d'évaluation
- A Historical Survey of Proposals To Transfer Palestinians From Palestine 1895 - 1947 - Transfer-Chaim-SimonsDocument297 pagesA Historical Survey of Proposals To Transfer Palestinians From Palestine 1895 - 1947 - Transfer-Chaim-SimonsBEN GURIONPas encore d'évaluation
- Congressaddresse00herz PDFDocument44 pagesCongressaddresse00herz PDFMoisés MotaPas encore d'évaluation
- 149064NCJRS PDFDocument47 pages149064NCJRS PDFMoisés Mota100% (2)
- Effects of Uoride On Rat Dental Enamel Matrix ProteinasesDocument8 pagesEffects of Uoride On Rat Dental Enamel Matrix ProteinasesMoisés MotaPas encore d'évaluation
- 12080924steuer - Zalomena OK1 PDFDocument21 pages12080924steuer - Zalomena OK1 PDFMoisés MotaPas encore d'évaluation
- The Spurious Rites of Memphis and Misraim - PIKE & CUMMINGS PDFDocument52 pagesThe Spurious Rites of Memphis and Misraim - PIKE & CUMMINGS PDFPierre Lalonde0% (1)
- Abbott Pharma Product ListingDocument103 pagesAbbott Pharma Product ListingNadeem AurangzebPas encore d'évaluation
- Sub Cloning Notebook EuroDocument72 pagesSub Cloning Notebook Eurod-fbuser-97323052Pas encore d'évaluation
- PCR Based Molecualr, Genetic MarkersDocument59 pagesPCR Based Molecualr, Genetic MarkersRafay MirzaPas encore d'évaluation
- PCR Detection of Bovine and Porcine Gelatin in Food ProductsDocument4 pagesPCR Detection of Bovine and Porcine Gelatin in Food ProductsAyten GüllücePas encore d'évaluation
- Hiv RapidDocument1 pageHiv RapidLutfi AhmadPas encore d'évaluation
- Case Analysis: Amgen Inc.: Pursuing Innovation AND IMITATION? (A) Case SolutionDocument2 pagesCase Analysis: Amgen Inc.: Pursuing Innovation AND IMITATION? (A) Case SolutionRicha Gupta0% (3)
- Identification of PTH-Amino Acids by HPLC: Gregory A. Grant and Mark W. CrankshawDocument22 pagesIdentification of PTH-Amino Acids by HPLC: Gregory A. Grant and Mark W. CrankshawJe RivasPas encore d'évaluation
- Cracraft & Donoghue (Eds.) - Assembling The Tree of Life (2004) PDFDocument594 pagesCracraft & Donoghue (Eds.) - Assembling The Tree of Life (2004) PDFAtman0% (1)
- ElisaDocument5 pagesElisaDennis ValdezPas encore d'évaluation
- The Update of The Phylogenetic Structure of Q1b Haplogroup Based On Full Y-Chromosome SequencingDocument9 pagesThe Update of The Phylogenetic Structure of Q1b Haplogroup Based On Full Y-Chromosome SequencingVladimir GurianovPas encore d'évaluation
- Proteinase K Manual 5prime 1044725 032007Document12 pagesProteinase K Manual 5prime 1044725 032007Irfan HussainPas encore d'évaluation
- Microbial Production of Scleroglucan and DSPDocument19 pagesMicrobial Production of Scleroglucan and DSPgandurikPas encore d'évaluation
- DNA Profiling or "Fingerprinting": Prepared By: Bersamina, Hanna Grace DDocument16 pagesDNA Profiling or "Fingerprinting": Prepared By: Bersamina, Hanna Grace DAlim Grace BersaminaPas encore d'évaluation
- Sanger Troubleshooting Guide v1Document5 pagesSanger Troubleshooting Guide v1yavuzPas encore d'évaluation
- List of Contributors - 2021 - Abiotic Stress and LegumesDocument6 pagesList of Contributors - 2021 - Abiotic Stress and LegumesPœta OciosoPas encore d'évaluation
- Preventing Rope SpoilageDocument2 pagesPreventing Rope SpoilageNhật Nguyễn SĩPas encore d'évaluation
- Cell Division Workbook - RemovedDocument6 pagesCell Division Workbook - RemovedSaurav YadavPas encore d'évaluation
- Tite Et Al 2019Document22 pagesTite Et Al 2019Jorge S. BurnsPas encore d'évaluation
- Food Chemistry: Analytical MethodsDocument6 pagesFood Chemistry: Analytical MethodsMihaelaOlaruPas encore d'évaluation
- Genetics and BiotechnologyDocument4 pagesGenetics and BiotechnologyCarla Itzel GtzPas encore d'évaluation
- Plant BiotechnologyDocument19 pagesPlant BiotechnologyZeeshan Ahmed100% (1)
- 5 Year Placement StatisticsDocument168 pages5 Year Placement StatisticsRam SskPas encore d'évaluation
- Curriculum Vitae: Date of Birth: 16th July, 1997 Place of Birth: Nayagarh, Odisha, India Citizenship: IndianDocument3 pagesCurriculum Vitae: Date of Birth: 16th July, 1997 Place of Birth: Nayagarh, Odisha, India Citizenship: IndianATMAJA ELINA MISHRAPas encore d'évaluation
- Microbiology Resource Announcements-2019-Saad-e00215-19.full - Grupo 5Document3 pagesMicrobiology Resource Announcements-2019-Saad-e00215-19.full - Grupo 5Wilgen Daniel Tapia CurillaPas encore d'évaluation
- Fermentation of Liquid BiofertilizersDocument5 pagesFermentation of Liquid BiofertilizersBrij Mohan SinghPas encore d'évaluation
- Melting Points of Organic ChemicalsDocument432 pagesMelting Points of Organic ChemicalsMAli RajaPas encore d'évaluation
- PfizerDocument10 pagesPfizerNayan ShuklaPas encore d'évaluation
- Residu Kanamycin HPLCDocument2 pagesResidu Kanamycin HPLCAprelita Nurelli Dwiana100% (1)
- JNTU KakinadaDocument5 pagesJNTU KakinadaAnweshaBosePas encore d'évaluation
- Biotech MCQs NumanDocument28 pagesBiotech MCQs NumansajjaddrPas encore d'évaluation