Vous aimerez peut-être aussi
- MIS DPCR IPCR July To December 2018 - Summary ListDocument14 pagesMIS DPCR IPCR July To December 2018 - Summary ListZyreen Kate BC100% (1)
- Work Permit Renewal Application FormDocument2 pagesWork Permit Renewal Application FormyamojodnabPas encore d'évaluation
- F.E. Campbell - Sweet Slavery - HIT 139Document115 pagesF.E. Campbell - Sweet Slavery - HIT 139HokusLocus67% (3)
- Blue Bloods 1Document166 pagesBlue Bloods 1anastomaPas encore d'évaluation
- Biosurfactant - Producing Bacte-Ria Found in Contaminated and Uncontaminated SoilsDocument8 pagesBiosurfactant - Producing Bacte-Ria Found in Contaminated and Uncontaminated SoilsYouly An RaidrisPas encore d'évaluation
- Melanized FungiDocument8 pagesMelanized FungiMuthu Vaduganathan NPas encore d'évaluation
- The Importance of Plants To Development and Maintenance of Soil StructureDocument11 pagesThe Importance of Plants To Development and Maintenance of Soil StructureRachel AzoubelPas encore d'évaluation
- Degradation of Paper Waste by Eisenia FoetidaDocument24 pagesDegradation of Paper Waste by Eisenia FoetidaAravind vishnu Saravanan GPas encore d'évaluation
- Aggelides 2000 PDFDocument7 pagesAggelides 2000 PDFjuan clarosPas encore d'évaluation
- 1-Soil Microbial Activity and Biomass As Influenced by Tillage and FertilizationDocument8 pages1-Soil Microbial Activity and Biomass As Influenced by Tillage and FertilizationTserendulam BatbaatarPas encore d'évaluation
- Agricultura Climáticamente Inteligente LibroDocument7 pagesAgricultura Climáticamente Inteligente LibroMarcela CuitivaPas encore d'évaluation
- TEXTO 1. Estudo de Caso (Dupla)Document5 pagesTEXTO 1. Estudo de Caso (Dupla)lucasgirioPas encore d'évaluation
- Biomassa e Atividade Microbiana Do Solo em Área de Cultivo de SojaDocument7 pagesBiomassa e Atividade Microbiana Do Solo em Área de Cultivo de SojaPaulo DiasPas encore d'évaluation
- Effect of Different Substrates On Vegetative Growth Characters of Tomato (Solanum Lycopersicum)Document8 pagesEffect of Different Substrates On Vegetative Growth Characters of Tomato (Solanum Lycopersicum)International Journal of Innovative Science and Research TechnologyPas encore d'évaluation
- SdarticleDocument8 pagesSdarticlejeffreygrajovelascoPas encore d'évaluation
- Earthworms and Vermitechnology A ReviewDocument12 pagesEarthworms and Vermitechnology A ReviewyyuzuakPas encore d'évaluation
- Bioresource Technology: Pritam Sangwan, C.P. Kaushik, V.K. GargDocument6 pagesBioresource Technology: Pritam Sangwan, C.P. Kaushik, V.K. Gargmudasirmalik011Pas encore d'évaluation
- Developing A Vegetable Fertility Program Using Organic Amendments and Inorganic FertilizersDocument8 pagesDeveloping A Vegetable Fertility Program Using Organic Amendments and Inorganic FertilizersDita Nidya KartikaPas encore d'évaluation
- Saxena 2001Document4 pagesSaxena 2001Kaelyn MontefalconPas encore d'évaluation
- VermivashDocument5 pagesVermivashb_manePas encore d'évaluation
- Overview of Earthworm Casts and A Comparison With CompostDocument24 pagesOverview of Earthworm Casts and A Comparison With CompostCk_psihPas encore d'évaluation
- Soil Factors Affect Fusarium Wilt SuppressionDocument8 pagesSoil Factors Affect Fusarium Wilt SuppressionClarisse MaximoPas encore d'évaluation
- Enhancement of Cotton Stalks Composting With Certain Microbial InoculationsDocument10 pagesEnhancement of Cotton Stalks Composting With Certain Microbial InoculationsKanhiya MahourPas encore d'évaluation
- Kastovska SBB2005Document9 pagesKastovska SBB2005Natasya Dwi ArthaMeliaPas encore d'évaluation
- 13750-Article Text-54096-1-10-20200601Document10 pages13750-Article Text-54096-1-10-20200601jonathan marcilloPas encore d'évaluation
- Sludge TreatmentDocument7 pagesSludge TreatmentRaluca-Cristina GuriencuPas encore d'évaluation
- 12524536Document5 pages12524536Shahriar ShamimPas encore d'évaluation
- Jurnal Indonesia: 11mu P ErtanianDocument7 pagesJurnal Indonesia: 11mu P ErtanianYuni RosianaPas encore d'évaluation
- 2007-Orlando Et Al SoilBiolBiochemDocument8 pages2007-Orlando Et Al SoilBiolBiochemJulieta OrlandoPas encore d'évaluation
- Research Introduction LACSINA - SALONGA 2 1Document9 pagesResearch Introduction LACSINA - SALONGA 2 1Lacsina QwyncyPas encore d'évaluation
- SdarticleDocument4 pagesSdarticleapi-3832223Pas encore d'évaluation
- Pinamonti 1998Document10 pagesPinamonti 1998Elisa DomínguezPas encore d'évaluation
- Perubahan Populasi Protozoa Dan Alga Dominan Pada Air Genangan Tanah Padi Sawah Yang Diberi Bokashi BerkelanjutanDocument7 pagesPerubahan Populasi Protozoa Dan Alga Dominan Pada Air Genangan Tanah Padi Sawah Yang Diberi Bokashi Berkelanjutannurul ummiPas encore d'évaluation
- Organic materials improve soil fertility in Liberica coffee landDocument10 pagesOrganic materials improve soil fertility in Liberica coffee landFalih WicaksonoPas encore d'évaluation
- Grellier, Riviere, Renault - 1999 - Transfer and Water-Retention Properties of Seed-Pelleting MaterialsDocument9 pagesGrellier, Riviere, Renault - 1999 - Transfer and Water-Retention Properties of Seed-Pelleting MaterialsJosé Matías ZapiolaPas encore d'évaluation
- Articulo Anita, AnitaDocument7 pagesArticulo Anita, AnitaAlejo MorenoPas encore d'évaluation
- Chamizo Et Al-2018-Frontiers in Environmental ScienceDocument14 pagesChamizo Et Al-2018-Frontiers in Environmental ScienceLucas Antonio GallaratoPas encore d'évaluation
- Bioresource Technology: Surindra SutharDocument7 pagesBioresource Technology: Surindra SutharSatawat TanaratPas encore d'évaluation
- Benefits and drawbacks of composting summarizedDocument6 pagesBenefits and drawbacks of composting summarizedRex ManserasPas encore d'évaluation
- Effect of Soil Solarization On Weed Seed Bank in SoilDocument2 pagesEffect of Soil Solarization On Weed Seed Bank in SoilSrujana PatelPas encore d'évaluation
- Seminar On Role of Organics in Maintenance of Soil HealthDocument29 pagesSeminar On Role of Organics in Maintenance of Soil HealthSimta SharmaPas encore d'évaluation
- Chapter 7-1 Gardening: Horticultural UsesDocument6 pagesChapter 7-1 Gardening: Horticultural UsesT CPas encore d'évaluation
- Dynamic Soil, Dynamic Plant Earthworms and Vermitechnology - A ReviewDocument13 pagesDynamic Soil, Dynamic Plant Earthworms and Vermitechnology - A ReviewDiego BautistaPas encore d'évaluation
- Studies On The Rhizosphere Mycoflora of MangrovesDocument5 pagesStudies On The Rhizosphere Mycoflora of MangrovesAdedayo A J AdewumiPas encore d'évaluation
- Bacterial and fungal taxon changes in soil microbial community composition induced by short-term biochar amendment in red oxidized loam soilDocument8 pagesBacterial and fungal taxon changes in soil microbial community composition induced by short-term biochar amendment in red oxidized loam soilFranky zPas encore d'évaluation
- Feeding Soil MacrofaunaDocument10 pagesFeeding Soil MacrofaunaNur Elva ZuhrahPas encore d'évaluation
- Organic Mulches Impact Soil Enzymes and EarthwormsDocument8 pagesOrganic Mulches Impact Soil Enzymes and EarthwormsBudi Mulyono100% (1)
- 17 IJAET Vol III Issue II 2012Document4 pages17 IJAET Vol III Issue II 2012gowthamiPas encore d'évaluation
- Effect of Vermicompost On Growth of Coconut Seedlings Under Field Conditions in Sri LankaDocument7 pagesEffect of Vermicompost On Growth of Coconut Seedlings Under Field Conditions in Sri LankaDave MuhammadPas encore d'évaluation
- Fungal communities in compost and vermicompostDocument12 pagesFungal communities in compost and vermicompostGabby VinuezaPas encore d'évaluation
- Comunidades Microbianas Del Suelo Bajo Sistemas de Cultivo y Cobertura Agroforestal de Cacao en El PerúDocument8 pagesComunidades Microbianas Del Suelo Bajo Sistemas de Cultivo y Cobertura Agroforestal de Cacao en El PerúERICA MERINOPas encore d'évaluation
- Journal of Environmental ManagementDocument8 pagesJournal of Environmental ManagementsadsdaPas encore d'évaluation
- Vermi CompostDocument102 pagesVermi CompostAnonymous L96u5itPas encore d'évaluation
- Informacion Sobre UvaDocument8 pagesInformacion Sobre UvaJesus Daniel Malpica GonzalezPas encore d'évaluation
- The State of Organic RightsDocument20 pagesThe State of Organic RightsmarielandryceoPas encore d'évaluation
- Protease Extraction From Soil by Sodium Pyrophosphate and Chemical Characterization of The ExtractsDocument13 pagesProtease Extraction From Soil by Sodium Pyrophosphate and Chemical Characterization of The ExtractsBassey NBPas encore d'évaluation
- 3 Kebede and Lele, 2022Document7 pages3 Kebede and Lele, 2022Tarekegn TeferaPas encore d'évaluation
- Isolation Methods for AzotobacterDocument8 pagesIsolation Methods for AzotobacterNarendra KumarPas encore d'évaluation
- Bioresource Technology: M.M. Coello-Castillo, J.E. Sánchez, D.J. RoyseDocument5 pagesBioresource Technology: M.M. Coello-Castillo, J.E. Sánchez, D.J. RoyseAbdelkader KhelifiPas encore d'évaluation
- Production of Oyster Mushroom On Different Substrates Using Cylindrical Block System R. C. Dey, K. M. Nasiruddin, M. S. Haque and M. A. Z. Al MunsurDocument6 pagesProduction of Oyster Mushroom On Different Substrates Using Cylindrical Block System R. C. Dey, K. M. Nasiruddin, M. S. Haque and M. A. Z. Al MunsurpilsamPas encore d'évaluation
- Growth and Yield Response of Marigold To Potting Media Containing Vermicompost Produced From Different WastesDocument9 pagesGrowth and Yield Response of Marigold To Potting Media Containing Vermicompost Produced From Different Wastespradhan.ayan23Pas encore d'évaluation
- Maintenance of Soil Functioning Following Erosion of Microbial DiversityDocument8 pagesMaintenance of Soil Functioning Following Erosion of Microbial DiversityAde Rizky FajrullohPas encore d'évaluation
- TMP EC3Document10 pagesTMP EC3FrontiersPas encore d'évaluation
- Communities and Ecosystems: Linking the Aboveground and Belowground Components (MPB-34)D'EverandCommunities and Ecosystems: Linking the Aboveground and Belowground Components (MPB-34)Pas encore d'évaluation
- Inside Out or Outside In. Meeting With Couples. Hugh JenkinsDocument24 pagesInside Out or Outside In. Meeting With Couples. Hugh JenkinsRita1976Pas encore d'évaluation
- Neuro EcDocument234 pagesNeuro EcFeynman2014100% (1)
- CH 2Document20 pagesCH 2Vivek SinghPas encore d'évaluation
- Wheat Sourdough Fermentation - Effects of Time and Acidification On Fundamental Rheological PropertiesDocument9 pagesWheat Sourdough Fermentation - Effects of Time and Acidification On Fundamental Rheological PropertiesmaurodiloretoPas encore d'évaluation
- Preparation of Starch Nanoparticles in A New Ionic Liquid-in-Oil MicroemulsionDocument12 pagesPreparation of Starch Nanoparticles in A New Ionic Liquid-in-Oil MicroemulsionPaula NancyPas encore d'évaluation
- WCDMA Optimization Case StudyDocument60 pagesWCDMA Optimization Case StudyRamesh Nikam100% (1)
- REMOTE Calculator Tool May 21 2013Document77 pagesREMOTE Calculator Tool May 21 2013Leyner Garcia MezquitaPas encore d'évaluation
- Schlumber: Kevin: 16010072 Moch Rizky Setiawan: 16010068Document12 pagesSchlumber: Kevin: 16010072 Moch Rizky Setiawan: 16010068KevinPas encore d'évaluation
- GRADES 1 To 12 Daily Lesson LogDocument5 pagesGRADES 1 To 12 Daily Lesson LogFlordeliza Manaois RamosPas encore d'évaluation
- Gaggia Cadorna Milk Full Instruction ManualDocument76 pagesGaggia Cadorna Milk Full Instruction ManualKanen Coffee, LLC.Pas encore d'évaluation
- An Experimental Study of AcidizingDocument11 pagesAn Experimental Study of Acidizinggigi17Pas encore d'évaluation
- How To Judge A Dasha in JyotishDocument2 pagesHow To Judge A Dasha in JyotishRavan SharmaPas encore d'évaluation
- Literature Review On Procrastination Impact On Academic PerformanceDocument2 pagesLiterature Review On Procrastination Impact On Academic Performancemomi100% (1)
- Sakshijain75.Final ProDocument46 pagesSakshijain75.Final ProOm AherPas encore d'évaluation
- Checklist PDFDocument84 pagesChecklist PDFJeremiah Miko LepasanaPas encore d'évaluation
- April 12th Test PDFDocument32 pagesApril 12th Test PDFهخه •Pas encore d'évaluation
- Periodic Test in Tle - Grade Ix Electricity.Document2 pagesPeriodic Test in Tle - Grade Ix Electricity.Resneth MondejarPas encore d'évaluation
- Effect of Non-Newtonian Fluids On The Performance of Plate Type Heat ExchangerDocument4 pagesEffect of Non-Newtonian Fluids On The Performance of Plate Type Heat Exchangerdeepika snehiPas encore d'évaluation
- Color Me VeganDocument272 pagesColor Me VeganBela Luísa Novo100% (1)
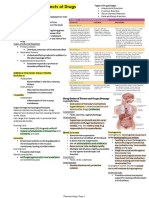
- CHAPTER 3: Toxic Effects of Drugs: Pharmacology Page 1Document1 pageCHAPTER 3: Toxic Effects of Drugs: Pharmacology Page 1Gabriel GonzagaPas encore d'évaluation
- Perfect Scale Pro ManualDocument9 pagesPerfect Scale Pro ManualAnthony VargasPas encore d'évaluation
- DILEM 10 G (24VDC) MoellerDocument7 pagesDILEM 10 G (24VDC) MoellerElizabeth RamirezPas encore d'évaluation
- Yoga & Diet To Prevent DepressionDocument5 pagesYoga & Diet To Prevent DepressionDietician Mrunmayee DixitPas encore d'évaluation
- Cat Helimax 2016 Esp 1Document74 pagesCat Helimax 2016 Esp 1HKM IngenierosPas encore d'évaluation
- RN and LPN Scope of Practice GuideDocument6 pagesRN and LPN Scope of Practice Guidejenn1722Pas encore d'évaluation
- Ramial Chipped Wood - A Basic Tool For Regenerating SoilsDocument15 pagesRamial Chipped Wood - A Basic Tool For Regenerating SoilsmrmiscPas encore d'évaluation