Vous aimerez peut-être aussi
- Species Concept, Speciation Evolutionary Thought Before DarwinDocument26 pagesSpecies Concept, Speciation Evolutionary Thought Before DarwinmashedtomatoezedPas encore d'évaluation
- Ethnobotany NotesDocument4 pagesEthnobotany NotesBereket Yitayaw100% (1)
- Animal DiversityDocument45 pagesAnimal DiversitySafiqa Tasfia100% (1)
- Introductory Entomology: Third Semester Lamjung CampusDocument206 pagesIntroductory Entomology: Third Semester Lamjung Campusnishan khatriPas encore d'évaluation
- CELL BILOGY AND GENETICS MANUAL (Practical 1 To 6)Document29 pagesCELL BILOGY AND GENETICS MANUAL (Practical 1 To 6)Ayesha FatimaPas encore d'évaluation
- Complete Chordata NotesDocument99 pagesComplete Chordata NotesNarasimha MurthyPas encore d'évaluation
- 06 GastrulationtxtDocument38 pages06 GastrulationtxtHafidzul HalimPas encore d'évaluation
- Ultrasonic Scaling TechniqueDocument3 pagesUltrasonic Scaling TechniquePutu WidiastriPas encore d'évaluation
- Extraembryonic MembranesDocument3 pagesExtraembryonic MembranesSimon RaoPas encore d'évaluation
- Plant Taxonomy NotesDocument38 pagesPlant Taxonomy Notesopolla nianorPas encore d'évaluation
- Chordate Origin and EvolutionDocument3 pagesChordate Origin and Evolutionmonique100% (2)
- Parasitism PDFDocument514 pagesParasitism PDFArvey Lozano100% (3)
- Lec4 SocialInsects TheSociety NotesDocument3 pagesLec4 SocialInsects TheSociety NotesAlexandra LvPas encore d'évaluation
- TOrsionDocument19 pagesTOrsionDrSneha VermaPas encore d'évaluation
- Drosophila MelanogasterDocument13 pagesDrosophila MelanogasterDavid MorganPas encore d'évaluation
- Theories of Organic EvolutionDocument15 pagesTheories of Organic EvolutionAdityaPas encore d'évaluation
- Phylum EchiuraDocument22 pagesPhylum EchiuraEthyl50% (2)
- Developmental Biology H. 277 - Gillbert BarresiDocument57 pagesDevelopmental Biology H. 277 - Gillbert BarresiAndini M RPas encore d'évaluation
- Plesiomorphy and SymplesiomorphyDocument3 pagesPlesiomorphy and SymplesiomorphyNTA UGC-NETPas encore d'évaluation
- Parasites & People - Host Parasite Relationship - RumalaDocument40 pagesParasites & People - Host Parasite Relationship - RumalamicroperadeniyaPas encore d'évaluation
- Genetic DriftDocument10 pagesGenetic DriftDimo PratannaPas encore d'évaluation
- Subkingdom Metazoa 2Document28 pagesSubkingdom Metazoa 2Ahmed OrabyPas encore d'évaluation
- Insects Growth and MetamorphosisDocument4 pagesInsects Growth and MetamorphosisAkash DashPas encore d'évaluation
- Sex Determination in DrosophilaDocument19 pagesSex Determination in DrosophilaSheerin Sulthana100% (2)
- Entomology Lab HandoutsDocument13 pagesEntomology Lab Handoutshumanupgrade100% (1)
- GENERAL BIOLOGY 4th Quarter - ACTIVITY #1-5Document7 pagesGENERAL BIOLOGY 4th Quarter - ACTIVITY #1-5Kimberly AyusoPas encore d'évaluation
- Evolution of PlantsDocument29 pagesEvolution of Plantstapas kunduPas encore d'évaluation
- Sampling and Isolation of Bacteria From SoilDocument9 pagesSampling and Isolation of Bacteria From SoilKishmala BatoolPas encore d'évaluation
- Metamorphism in InsectsDocument24 pagesMetamorphism in Insectsyayeg raja100% (1)
- Cnidaria: Body Cavity Known As The CoelenteronDocument21 pagesCnidaria: Body Cavity Known As The CoelenteronLakshya GuptaPas encore d'évaluation
- Entamoeba HistolyticaDocument18 pagesEntamoeba HistolyticaMayuri VohraPas encore d'évaluation
- 2010 MG EntomologyDocument11 pages2010 MG EntomologyhundohanPas encore d'évaluation
- Whitefly Biology PDFDocument27 pagesWhitefly Biology PDFJulian LeonardoPas encore d'évaluation
- Parasitology NotesDocument3 pagesParasitology NotesMyrielle Trisha SAYREPas encore d'évaluation
- 07 Gastrulation BiologyDocument22 pages07 Gastrulation BiologyKashif JamalPas encore d'évaluation
- Assignment 1589713070 SmsDocument4 pagesAssignment 1589713070 SmsSuchitra SharmaPas encore d'évaluation
- Phylum Echinodermata: Starfish, Sea Urchin, Sea Brittle, Sea Lilies, Sea Cucumber, Sea FeatherDocument31 pagesPhylum Echinodermata: Starfish, Sea Urchin, Sea Brittle, Sea Lilies, Sea Cucumber, Sea FeatherAmmy SyazannaPas encore d'évaluation
- History of Insect ClassificationDocument11 pagesHistory of Insect Classificationapi-685744638Pas encore d'évaluation
- Cytoplasmic InheritanceDocument13 pagesCytoplasmic InheritanceIzza NafisaPas encore d'évaluation
- Bio102 General Biology II SummaryDocument47 pagesBio102 General Biology II SummaryIkenna Okpala100% (1)
- Unit: 08 Placentation in Mammals: Developmental Biology Cr. Hours: 4 (3+1)Document30 pagesUnit: 08 Placentation in Mammals: Developmental Biology Cr. Hours: 4 (3+1)Task BirdPas encore d'évaluation
- Echinoderm Larvae PDFDocument15 pagesEchinoderm Larvae PDFMadhav GoelPas encore d'évaluation
- Evolution Review Packet Key EditableDocument4 pagesEvolution Review Packet Key EditableMiljoy Delegado100% (1)
- 4+5 Tissue Types and Epithelial TissueDocument44 pages4+5 Tissue Types and Epithelial Tissueyasser100% (2)
- Animal Form and Function: C E L PDocument38 pagesAnimal Form and Function: C E L PJoshua Miguel Buno0% (1)
- Phylum PoriferaDocument17 pagesPhylum PoriferaRhiZhal SquDetto FrizTalPas encore d'évaluation
- Systematic Approaches To Phylogeny)Document26 pagesSystematic Approaches To Phylogeny)Mhi IsmailPas encore d'évaluation
- M - 23 Polytene ChromosomeDocument3 pagesM - 23 Polytene ChromosomeDr. Tapan Kr. DuttaPas encore d'évaluation
- Biology Cls 12 ProjectDocument13 pagesBiology Cls 12 ProjectARITRA KUNDUPas encore d'évaluation
- Insect Reproduction CK Dash SAUDocument3 pagesInsect Reproduction CK Dash SAUChandra Kanta Dash100% (1)
- AlgaeDocument34 pagesAlgaeAnand KumarPas encore d'évaluation
- Nematode NotesDocument2 pagesNematode Notesapi-247084136100% (1)
- Classification of PoriferaDocument14 pagesClassification of PoriferaHastin Atas AsihPas encore d'évaluation
- Animal Diversity 1 PDFDocument20 pagesAnimal Diversity 1 PDFRica Jane Delan MapaloPas encore d'évaluation
- Plant Viruses As Molecular PathogensDocument557 pagesPlant Viruses As Molecular PathogensDare SpartuPas encore d'évaluation
- 2Document3 pages2Kyrish DaceraPas encore d'évaluation
- BryophytesDocument12 pagesBryophytesAiman KaleemPas encore d'évaluation
- Recombinant DNA And Cell ProliferationD'EverandRecombinant DNA And Cell ProliferationGary SteinPas encore d'évaluation
- TF 22645580Document4 pagesTF 22645580Sree NaveenaPas encore d'évaluation
- Presentation1 M.SC EntoDocument25 pagesPresentation1 M.SC EntoSree NaveenaPas encore d'évaluation
- Presentation1 M.SC EntoDocument25 pagesPresentation1 M.SC EntoSree NaveenaPas encore d'évaluation
- PAT 577 Assignment On Diseases of BajraDocument17 pagesPAT 577 Assignment On Diseases of BajraSree Naveena100% (1)
- PAT 577 Assignment On Diseases of BajraDocument17 pagesPAT 577 Assignment On Diseases of BajraSree Naveena100% (1)
- Sawit - INSECT POLLINATION BY INTRODUCTION OF ELAEIDOBIUS KAMERUNICUSDocument2 pagesSawit - INSECT POLLINATION BY INTRODUCTION OF ELAEIDOBIUS KAMERUNICUSJob Zine'dine100% (1)
- Warbiter: Oak Bush CricketDocument11 pagesWarbiter: Oak Bush Cricketpaulus gunardi agungPas encore d'évaluation
- Insect Wings, Its Venation, Modifications and Wing Coupling: Ravy RaazDocument11 pagesInsect Wings, Its Venation, Modifications and Wing Coupling: Ravy RaazParvendra Yadav YadavPas encore d'évaluation
- Bee Life Cycle 1Document6 pagesBee Life Cycle 1Nidhi ChandraPas encore d'évaluation
- Insect AriumDocument11 pagesInsect AriumBabasChongPas encore d'évaluation
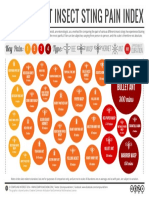
- The Schmidt Insect Sting Pain Index PDFDocument1 pageThe Schmidt Insect Sting Pain Index PDFDaniel MorenaPas encore d'évaluation
- Vectors of Public Health ImportanceDocument31 pagesVectors of Public Health ImportanceJoseph Stans Kasirye75% (4)
- OdonataDocument12 pagesOdonataJuste NavickaitePas encore d'évaluation
- Family: Hippoboscidae: Common Name: Pupipara, Forest FliesDocument28 pagesFamily: Hippoboscidae: Common Name: Pupipara, Forest FliesAvitus Dastan CastoPas encore d'évaluation
- Butterfly Life Cycle With ScriptDocument22 pagesButterfly Life Cycle With ScriptJuju GarlanPas encore d'évaluation
- Flight of the Bumblebee: Piano Arr By Rachmaninoff 製譜 By Nice Piano SheetsDocument6 pagesFlight of the Bumblebee: Piano Arr By Rachmaninoff 製譜 By Nice Piano SheetslaraloPas encore d'évaluation
- 2 MosquitoesDocument41 pages2 MosquitoesAnmut AschalePas encore d'évaluation
- Paper Wasps: General InformationDocument1 pagePaper Wasps: General InformationLinesh BrahmakhatriPas encore d'évaluation
- Caracterisiticas Basicas de La Hermetia IllucensDocument2 pagesCaracterisiticas Basicas de La Hermetia IllucensalbperPas encore d'évaluation
- Musca and StomoxysDocument8 pagesMusca and StomoxysransinghPas encore d'évaluation
- Crop Protection 1Document22 pagesCrop Protection 1BONGO John Ryan C.Pas encore d'évaluation
- Wee Sing and Learn Bugs: (Song Lyrics) Bugs, BugsDocument5 pagesWee Sing and Learn Bugs: (Song Lyrics) Bugs, BugsCsilla SzabóPas encore d'évaluation
- LP 5 Clasificarea Insectelor I - ImaginiDocument7 pagesLP 5 Clasificarea Insectelor I - ImaginiMadalina VasilePas encore d'évaluation
- All Aluminium Conductors AAC Bs 215 Part 1Document1 pageAll Aluminium Conductors AAC Bs 215 Part 1nigerluvPas encore d'évaluation
- Feeding Process and Mouth Parts of InsectDocument7 pagesFeeding Process and Mouth Parts of InsectTayyabPas encore d'évaluation
- Ants IdentificationDocument63 pagesAnts IdentificationGelu TiberiuPas encore d'évaluation
- 4.dermestid BeetleDocument2 pages4.dermestid Beetlevandv printsPas encore d'évaluation
- Tle-10 Afa Oa q3 Las 1Document13 pagesTle-10 Afa Oa q3 Las 1Yves Kim Casido CabantingPas encore d'évaluation
- BSC Agriculture 3 Sem Fundamentals of Insect Morphology and Systematics 72553 May 2019Document2 pagesBSC Agriculture 3 Sem Fundamentals of Insect Morphology and Systematics 72553 May 2019Navneet KumarPas encore d'évaluation
- Learn ABout Texas InsectsDocument64 pagesLearn ABout Texas Insectsdaiana_olteanuPas encore d'évaluation
- Insects HeadDocument33 pagesInsects HeadRavi,50% (2)
- 4250 Lec18 15Document13 pages4250 Lec18 15Carlos Enrique Pijo PerezPas encore d'évaluation
- The Night Moth With A Crooked FeelerDocument2 pagesThe Night Moth With A Crooked FeelerLaura ArauzPas encore d'évaluation
- 3rd Social InsectsDocument2 pages3rd Social InsectsyvhonnedelimaPas encore d'évaluation
- LS9 9Document1 pageLS9 9sinned68Pas encore d'évaluation