Vous aimerez peut-être aussi
- Food Control: Peter M. Horchner, Andrew M. PointonDocument15 pagesFood Control: Peter M. Horchner, Andrew M. PointoncikizdPas encore d'évaluation
- Chapter One 1.1 Background To The StudyDocument6 pagesChapter One 1.1 Background To The StudyIgbani VictoryPas encore d'évaluation
- Basic Hygiene Practice in Meat IndustryDocument3 pagesBasic Hygiene Practice in Meat IndustryZaheer HussainPas encore d'évaluation
- Contribution of Meat Inspection To The Surveillance of Poultry Health and Welfare in The European UnionDocument14 pagesContribution of Meat Inspection To The Surveillance of Poultry Health and Welfare in The European UnionShah NawazPas encore d'évaluation
- 24بحث رقم 4421 عدد يوليو 2013Document11 pages24بحث رقم 4421 عدد يوليو 2013ابراهيم القويعىPas encore d'évaluation
- Meat Milk and Fish HygieneDocument74 pagesMeat Milk and Fish Hygieneransingh100% (1)
- Microbial Decontamination, Food Safety, and Antimicrobial InterventionsDocument6 pagesMicrobial Decontamination, Food Safety, and Antimicrobial InterventionsZeyar Than WinPas encore d'évaluation
- Importance of Quality Assurance For Livestock TradeDocument7 pagesImportance of Quality Assurance For Livestock TradeBharat MundhePas encore d'évaluation
- Food Hygiene:: Escherichia Coli O157:H7 and The Infectious Agent of Bovine/transmissible SpongiformDocument4 pagesFood Hygiene:: Escherichia Coli O157:H7 and The Infectious Agent of Bovine/transmissible SpongiformXher FaixalPas encore d'évaluation
- Field Report Update 2023Document21 pagesField Report Update 2023Eyoseyas GetachewPas encore d'évaluation
- Ai 410 e 00Document27 pagesAi 410 e 00Lukman PitonoPas encore d'évaluation
- Abattoir DevelopmentDocument101 pagesAbattoir Developmentreenasaini21097713100% (4)
- Food SafetyDocument3 pagesFood SafetyFederico LeonPas encore d'évaluation
- Major Causes of Organ and Carcass Condemnation in Cattle Slaughtered at Bahir Dar Municipality Abattoir, North West EthiopiaDocument9 pagesMajor Causes of Organ and Carcass Condemnation in Cattle Slaughtered at Bahir Dar Municipality Abattoir, North West EthiopiaPremier PublishersPas encore d'évaluation
- Dorathy Original (2) ProjectDocument32 pagesDorathy Original (2) ProjectEMMANUEL SALVATIONPas encore d'évaluation
- A Review of Shiga-Toxin Producing Escherichia Coli (STEC) Contamination in The Raw Pork Production ChainDocument8 pagesA Review of Shiga-Toxin Producing Escherichia Coli (STEC) Contamination in The Raw Pork Production ChainHtet Lin OoPas encore d'évaluation
- Isolation and Molecular Characterization of Salmonella Enterica, Escherichia Coli O157-H7 and Shigella Spp. From Meat and Dairy Products in EgyptDocument6 pagesIsolation and Molecular Characterization of Salmonella Enterica, Escherichia Coli O157-H7 and Shigella Spp. From Meat and Dairy Products in EgyptAffan GaffarPas encore d'évaluation
- Assessment of Hygienic Practice On Camel Meat Handlers, and Identification of Main Points of Bacterial Contamination in Abattoir and Butcheries of Nagelle Town, Southern Oromia, EthiopiaDocument17 pagesAssessment of Hygienic Practice On Camel Meat Handlers, and Identification of Main Points of Bacterial Contamination in Abattoir and Butcheries of Nagelle Town, Southern Oromia, Ethiopiaresearchparks100% (1)
- Food Safety Issues Affecting The Dairy Beef Industry: Gary StefanDocument5 pagesFood Safety Issues Affecting The Dairy Beef Industry: Gary Stefancomercial amcPas encore d'évaluation
- Efsa Riscos em Carne de Aves 2012Document179 pagesEfsa Riscos em Carne de Aves 2012alberto.afonso3748Pas encore d'évaluation
- Food Safety: Fact SheetsDocument2 pagesFood Safety: Fact Sheetslox agencyPas encore d'évaluation
- Journal of The Science of Food and Agriculture 2017 97 (4) 1073-1083Document11 pagesJournal of The Science of Food and Agriculture 2017 97 (4) 1073-1083Kobi ShimshoniPas encore d'évaluation
- Chemical and Microbial Assessment of BeeDocument9 pagesChemical and Microbial Assessment of BeeDaviid A'SPas encore d'évaluation
- Nwanta Et AlDocument8 pagesNwanta Et AlJoko LumawagPas encore d'évaluation
- DescargaDocument7 pagesDescargaRicardo QuimisPas encore d'évaluation
- REFERAT - Salinomycin Intoxication in TurkeysDocument4 pagesREFERAT - Salinomycin Intoxication in TurkeysCotovici Catana AlinaPas encore d'évaluation
- Executive Summary: Supporting Document 4Document8 pagesExecutive Summary: Supporting Document 4ghajiniiPas encore d'évaluation
- Jurnal Acuan 2Document14 pagesJurnal Acuan 2Dwi IryantiPas encore d'évaluation
- Ivan NastasijevicDocument15 pagesIvan NastasijevicmiskoscribdPas encore d'évaluation
- Meat Products in The Past, Today and in The FutureDocument10 pagesMeat Products in The Past, Today and in The FuturedknwPas encore d'évaluation
- Food Safety, Labeling .AshxDocument36 pagesFood Safety, Labeling .AshxFalguniPas encore d'évaluation
- Eje Cafetero BienestarDocument8 pagesEje Cafetero BienestarAlejandra Gomez RuizPas encore d'évaluation
- A HACCP Plan For Mycotoxigenic Hazards Associated With Dry-Cured Meat Production ProcessesDocument7 pagesA HACCP Plan For Mycotoxigenic Hazards Associated With Dry-Cured Meat Production ProcesseshilalpunooPas encore d'évaluation
- Foodbalt 2014 Microbiological Quality of Meat Preparations and Meat ProductsDocument5 pagesFoodbalt 2014 Microbiological Quality of Meat Preparations and Meat ProductsRagil PamungkasPas encore d'évaluation
- Working Conditions in Serowe Regional LandfillDocument12 pagesWorking Conditions in Serowe Regional LandfillEMMANUEL BOTSHELOPas encore d'évaluation
- Foods 09 00546Document10 pagesFoods 09 00546armaan aliPas encore d'évaluation
- BIOHAZ PiensosSalmonella08Document84 pagesBIOHAZ PiensosSalmonella08amamùra maamarPas encore d'évaluation
- EFSA Journal - 2012Document179 pagesEFSA Journal - 2012Jiagen GongPas encore d'évaluation
- Haccp For Irish Beef, Pork and Lamb Slaughter: AuthorsDocument16 pagesHaccp For Irish Beef, Pork and Lamb Slaughter: Authorspopa.janossPas encore d'évaluation
- Policy EditedDocument57 pagesPolicy EditedJoann SacolPas encore d'évaluation
- Good Practices in Primary Production: Section 2Document22 pagesGood Practices in Primary Production: Section 2Nilamdeen Mohamed ZamilPas encore d'évaluation
- Trazabilida VerDocument14 pagesTrazabilida Verabraham84Pas encore d'évaluation
- Meat Science: C.E. Realini, M. Font I Furnols, C. Sañudo, F. Montossi, M.A. Oliver, L. GuerreroDocument8 pagesMeat Science: C.E. Realini, M. Font I Furnols, C. Sañudo, F. Montossi, M.A. Oliver, L. GuerreroAndreea MarinPas encore d'évaluation
- MicrobiologicalDocument12 pagesMicrobiologicalTrung Truong NhatPas encore d'évaluation
- Report-Slaughter Workshop - Richard - Klottey - BotchwayDocument11 pagesReport-Slaughter Workshop - Richard - Klottey - BotchwayRichard Klottey BotchwayPas encore d'évaluation
- Beuc-X-2014-052 Cpe Beuc Position Paper-Use of Peroxyacetic Acid On Poultry Carcases and MeatDocument11 pagesBeuc-X-2014-052 Cpe Beuc Position Paper-Use of Peroxyacetic Acid On Poultry Carcases and MeatMohamed RayPas encore d'évaluation
- Poultry - Abattoir Good Practices ThailandDocument22 pagesPoultry - Abattoir Good Practices ThailandJaime SilvaPas encore d'évaluation
- Study The Presence of Escherichia Coli in Broiler Chicken Meat As Potential Public Health ThreatDocument4 pagesStudy The Presence of Escherichia Coli in Broiler Chicken Meat As Potential Public Health ThreatHuma AhmadyPas encore d'évaluation
- Isolation SamonellaDocument8 pagesIsolation SamonellaHoai LePas encore d'évaluation
- Duc 2018Document8 pagesDuc 2018priscillaPas encore d'évaluation
- Guidance Note Meat Poultry 03-02-2021Document10 pagesGuidance Note Meat Poultry 03-02-2021NATTY GAINSPas encore d'évaluation
- Literature Review On Meat HygieneDocument6 pagesLiterature Review On Meat Hygieneekvxrxrif100% (1)
- GMP Dry SausageDocument26 pagesGMP Dry SausageJade NguyenPas encore d'évaluation
- Identification of Cattle Poisoning by Bifenthrin Via EarwaxDocument24 pagesIdentification of Cattle Poisoning by Bifenthrin Via EarwaxNAYARA MOREIRA DAMASCENOPas encore d'évaluation
- Application of Haccp Principles in The Meat Industry: United Kingdom PerspectiveDocument9 pagesApplication of Haccp Principles in The Meat Industry: United Kingdom PerspectiveBianca IonescuPas encore d'évaluation
- Antimicrobial ResidueDocument10 pagesAntimicrobial ResidueGigiPetreaPas encore d'évaluation
- The Veterinarian's Role in Food SafetyDocument16 pagesThe Veterinarian's Role in Food SafetyakdasivriPas encore d'évaluation
- Benefits of Routine Biosecurity Measures in Commercial Layer FarmsDocument9 pagesBenefits of Routine Biosecurity Measures in Commercial Layer FarmsGabrielPas encore d'évaluation
- 7.microbiology of MeatsDocument21 pages7.microbiology of MeatsMahesh DevasigamaniPas encore d'évaluation
- Sample Haad QuestionDocument15 pagesSample Haad QuestionFaith Levi Alecha AlferezPas encore d'évaluation
- Drug StudyDocument8 pagesDrug StudyAmsh RosalPas encore d'évaluation
- Aging and Down Syndrome A Health and Well-Being GuidebookDocument44 pagesAging and Down Syndrome A Health and Well-Being GuidebookNicolePas encore d'évaluation
- Niosome Final...Document9 pagesNiosome Final...Ajma LinePas encore d'évaluation
- Acute Abdominal PainDocument5 pagesAcute Abdominal PainLM Mys100% (1)
- Effect of Ethanol Leaf Extract of Chromolaena Odorata On Lipid Profile of Streptozotocin Induced Diabetic Wistar Albino RatsDocument9 pagesEffect of Ethanol Leaf Extract of Chromolaena Odorata On Lipid Profile of Streptozotocin Induced Diabetic Wistar Albino RatsPUBLISHER JOURNALPas encore d'évaluation
- Guidelines For Sedation and Anesthesia in GI Endos PDFDocument11 pagesGuidelines For Sedation and Anesthesia in GI Endos PDFHernan RuedaPas encore d'évaluation
- Respiratory Physiology Lab ReportDocument15 pagesRespiratory Physiology Lab ReportThalia PacamalanPas encore d'évaluation
- Gyne NCPDocument3 pagesGyne NCPJoshua ArevaloPas encore d'évaluation
- Osteoarthritis of The KneeDocument634 pagesOsteoarthritis of The KneeSergiu Plesca100% (2)
- Pi Is 0025619616308187Document13 pagesPi Is 0025619616308187Anthony M CaballeroPas encore d'évaluation
- The Client With Chronic Renal FailureDocument4 pagesThe Client With Chronic Renal FailureMarisol Jane JomayaPas encore d'évaluation
- Repertory 210313 SyllabusDocument6 pagesRepertory 210313 SyllabusamoldholePas encore d'évaluation
- Iaea TRS 469 PDFDocument86 pagesIaea TRS 469 PDFClaudia Morales UlloaPas encore d'évaluation
- JONAM16000170Document14 pagesJONAM16000170R.G.ManikarajanPas encore d'évaluation
- Collecting A Pediatric Urine SpecimenDocument2 pagesCollecting A Pediatric Urine SpecimenLarry CornejoPas encore d'évaluation
- Improving The Quality of Health Journalism: When Reliability Meets Engagement, Heini MaksimainenDocument60 pagesImproving The Quality of Health Journalism: When Reliability Meets Engagement, Heini MaksimainenRed Ética Segura - FNPIPas encore d'évaluation
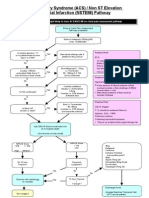
- ACS NSTEMI Clinical PathwayDocument3 pagesACS NSTEMI Clinical PathwayXtiaRPas encore d'évaluation
- Intentional Living Worksheet - Copyright - Day Designer - Do Not Distribute - 03-17 PDFDocument1 pageIntentional Living Worksheet - Copyright - Day Designer - Do Not Distribute - 03-17 PDFCourtney JohnsonPas encore d'évaluation
- Patty NCP HyperthermiaDocument4 pagesPatty NCP HyperthermiaPatricia Jean FaeldoneaPas encore d'évaluation
- Sindh Nurses Examination Board Karachi: Center Supervisor: S.No Name School Name RemarksDocument1 pageSindh Nurses Examination Board Karachi: Center Supervisor: S.No Name School Name RemarksAyazPas encore d'évaluation
- UW ObjectivesDocument171 pagesUW ObjectivesCarolina Lopez0% (1)
- Clinical and Etiological Profile of Acute Febrile Encephalopathy in Eastern NepalDocument3 pagesClinical and Etiological Profile of Acute Febrile Encephalopathy in Eastern NepalNeha OberoiPas encore d'évaluation
- JBDS 02 Nursing Management For DKA v1 20122022Document10 pagesJBDS 02 Nursing Management For DKA v1 20122022olazbt2000Pas encore d'évaluation
- Poster PPT OssnDocument12 pagesPoster PPT OssnShilpa B KPas encore d'évaluation
- Antipsychotic DrugsDocument3 pagesAntipsychotic DrugsArtt1975Pas encore d'évaluation
- Bmi Data May Joie J. AlcarazDocument13 pagesBmi Data May Joie J. AlcarazMay Joie JaymePas encore d'évaluation
- 16-Clinical Anatomy of The Upper LimbDocument27 pages16-Clinical Anatomy of The Upper LimbAlapati Vinod KumarPas encore d'évaluation
- Yellow Card 20 MedicDocument2 pagesYellow Card 20 MedicLeonardo AlcotaPas encore d'évaluation
- An Introduction To MHRA GCP Inspections - : Michelle Rowson, GCP InspectorDocument26 pagesAn Introduction To MHRA GCP Inspections - : Michelle Rowson, GCP InspectorSwapnil PanpatilPas encore d'évaluation
- The Diabetes Code: Prevent and Reverse Type 2 Diabetes NaturallyD'EverandThe Diabetes Code: Prevent and Reverse Type 2 Diabetes NaturallyÉvaluation : 4.5 sur 5 étoiles4.5/5 (4)
- The Diet Trap Solution: Train Your Brain to Lose Weight and Keep It Off for GoodD'EverandThe Diet Trap Solution: Train Your Brain to Lose Weight and Keep It Off for GoodPas encore d'évaluation
- Summary of Mary Claire Haver's The Galveston DietD'EverandSummary of Mary Claire Haver's The Galveston DietÉvaluation : 5 sur 5 étoiles5/5 (2)
- The Candida Cure: The 90-Day Program to Balance Your Gut, Beat Candida, and Restore Vibrant HealthD'EverandThe Candida Cure: The 90-Day Program to Balance Your Gut, Beat Candida, and Restore Vibrant HealthPas encore d'évaluation
- Find Your Path: Honor Your Body, Fuel Your Soul, and Get Strong with the Fit52 LifeD'EverandFind Your Path: Honor Your Body, Fuel Your Soul, and Get Strong with the Fit52 LifeÉvaluation : 4 sur 5 étoiles4/5 (3)
- Sugar Crush: How to Reduce Inflammation, Reverse Nerve Damage, and Reclaim Good HealthD'EverandSugar Crush: How to Reduce Inflammation, Reverse Nerve Damage, and Reclaim Good HealthÉvaluation : 4 sur 5 étoiles4/5 (6)
- The Noom Kitchen: 100 Healthy, Delicious, Flexible Recipes for Every DayD'EverandThe Noom Kitchen: 100 Healthy, Delicious, Flexible Recipes for Every DayPas encore d'évaluation
- Forever Strong: A New, Science-Based Strategy for Aging WellD'EverandForever Strong: A New, Science-Based Strategy for Aging WellPas encore d'évaluation
- The Beck Diet Solution Weight Loss Workbook: The 6-Week Plan to Train Your Brain to Think Like a Thin PersonD'EverandThe Beck Diet Solution Weight Loss Workbook: The 6-Week Plan to Train Your Brain to Think Like a Thin PersonÉvaluation : 3.5 sur 5 étoiles3.5/5 (33)
- Foods That Cause You to Lose Weight: The Negative Calorie EffectD'EverandFoods That Cause You to Lose Weight: The Negative Calorie EffectÉvaluation : 3 sur 5 étoiles3/5 (5)
- The Arm: Inside the Billion-Dollar Mystery of the Most Valuable Commodity in SportsD'EverandThe Arm: Inside the Billion-Dollar Mystery of the Most Valuable Commodity in SportsÉvaluation : 4 sur 5 étoiles4/5 (49)
- Secrets From the Eating Lab: The Science of Weight Loss, the Myth of Willpower, and Why You Should Never Diet AgainD'EverandSecrets From the Eating Lab: The Science of Weight Loss, the Myth of Willpower, and Why You Should Never Diet AgainÉvaluation : 3.5 sur 5 étoiles3.5/5 (38)
- Metabolism Revolution: Lose 14 Pounds in 14 Days and Keep It Off for LifeD'EverandMetabolism Revolution: Lose 14 Pounds in 14 Days and Keep It Off for LifePas encore d'évaluation
- Glucose Revolution: The Life-Changing Power of Balancing Your Blood SugarD'EverandGlucose Revolution: The Life-Changing Power of Balancing Your Blood SugarÉvaluation : 5 sur 5 étoiles5/5 (352)
- Body Love Every Day: Choose Your Life-Changing 21-Day Path to Food FreedomD'EverandBody Love Every Day: Choose Your Life-Changing 21-Day Path to Food FreedomÉvaluation : 4 sur 5 étoiles4/5 (1)
- Eat & Run: My Unlikely Journey to Ultramarathon GreatnessD'EverandEat & Run: My Unlikely Journey to Ultramarathon GreatnessPas encore d'évaluation
- The Body Book: The Law of Hunger, the Science of Strength, and Other Ways to Love Your Amazing BodyD'EverandThe Body Book: The Law of Hunger, the Science of Strength, and Other Ways to Love Your Amazing BodyPas encore d'évaluation
- Grit & Grace: Train the Mind, Train the Body, Own Your LifeD'EverandGrit & Grace: Train the Mind, Train the Body, Own Your LifeÉvaluation : 4 sur 5 étoiles4/5 (3)
- Instant Loss On a Budget: Super-Affordable Recipes for the Health-Conscious CookD'EverandInstant Loss On a Budget: Super-Affordable Recipes for the Health-Conscious CookÉvaluation : 3.5 sur 5 étoiles3.5/5 (2)
- Hungry for Change: Ditch the Diets, Conquer the Cravings, and Eat Your Way to Lifelong HealthD'EverandHungry for Change: Ditch the Diets, Conquer the Cravings, and Eat Your Way to Lifelong HealthÉvaluation : 4 sur 5 étoiles4/5 (7)
- Eat to Lose, Eat to Win: Your Grab-n-Go Action Plan for a Slimmer, Healthier YouD'EverandEat to Lose, Eat to Win: Your Grab-n-Go Action Plan for a Slimmer, Healthier YouPas encore d'évaluation
- The UltraMind Solution: Fix Your Broken Brain by Healing Your Body FirstD'EverandThe UltraMind Solution: Fix Your Broken Brain by Healing Your Body FirstÉvaluation : 4.5 sur 5 étoiles4.5/5 (49)
- The Food Lover's Cleanse: 140 Delicious, Nourishing Recipes That Will Tempt You Back into Healthful EatingD'EverandThe Food Lover's Cleanse: 140 Delicious, Nourishing Recipes That Will Tempt You Back into Healthful EatingÉvaluation : 4 sur 5 étoiles4/5 (3)
- The Volumetrics Eating Plan: Techniques and Recipes for Feeling Full on Fewer CaloriesD'EverandThe Volumetrics Eating Plan: Techniques and Recipes for Feeling Full on Fewer CaloriesPas encore d'évaluation
- How to Be Well: The 6 Keys to a Happy and Healthy LifeD'EverandHow to Be Well: The 6 Keys to a Happy and Healthy LifeÉvaluation : 5 sur 5 étoiles5/5 (1)
- Glucose Goddess Method: A 4-Week Guide to Cutting Cravings, Getting Your Energy Back, and Feeling AmazingD'EverandGlucose Goddess Method: A 4-Week Guide to Cutting Cravings, Getting Your Energy Back, and Feeling AmazingÉvaluation : 5 sur 5 étoiles5/5 (61)
- Rapid Weight Loss Hypnosis: Deep Sleep Your Way to Rapid Weight Loss, Healing Your Body and Self Esteem with Guided Meditations and Positive AffirmationsD'EverandRapid Weight Loss Hypnosis: Deep Sleep Your Way to Rapid Weight Loss, Healing Your Body and Self Esteem with Guided Meditations and Positive AffirmationsÉvaluation : 5 sur 5 étoiles5/5 (7)