Vous aimerez peut-être aussi
- Effect of Immunozin On Sickle Cell Disease in Sub Saharan Africa A Pilot StudyDocument11 pagesEffect of Immunozin On Sickle Cell Disease in Sub Saharan Africa A Pilot StudymohammedPas encore d'évaluation
- Jiw 427Document8 pagesJiw 427Anderson LondoñoPas encore d'évaluation
- Sickle Cell DiseaseDocument10 pagesSickle Cell Diseasekarthikavvari2002Pas encore d'évaluation
- Thalassemias Ali T Taher, David J Weatherall, Maria Domenica Cappellini 2018Document13 pagesThalassemias Ali T Taher, David J Weatherall, Maria Domenica Cappellini 2018Malika MohPas encore d'évaluation
- A New Step in The Treatment of Sickle Cell DiseaseDocument2 pagesA New Step in The Treatment of Sickle Cell Disease422073224Pas encore d'évaluation
- Leptospirosis CasosDocument2 pagesLeptospirosis CasosfelipePas encore d'évaluation
- Research ArticleDocument7 pagesResearch ArticleerikaPas encore d'évaluation
- Sickle Cell Disease: Advances in TreatmentDocument13 pagesSickle Cell Disease: Advances in TreatmentarturomarticarvajalPas encore d'évaluation
- Malaria and The Red CellDocument23 pagesMalaria and The Red CellLinda FatrisiaPas encore d'évaluation
- Annotated en - 0120 9957 RCG 34 04 00433Document4 pagesAnnotated en - 0120 9957 RCG 34 04 00433Gguk yemPas encore d'évaluation
- Artigo - Gitelman Syndrome ÍntegraDocument2 pagesArtigo - Gitelman Syndrome Íntegrasalvianolucas5Pas encore d'évaluation
- qt4c13q7gw PDFDocument19 pagesqt4c13q7gw PDFAb Al-mamariPas encore d'évaluation
- Seminar: Ali T Taher, David J Weatherall, Maria Domenica CappelliniDocument13 pagesSeminar: Ali T Taher, David J Weatherall, Maria Domenica CappelliniCarlos MorenoPas encore d'évaluation
- Hematology 2006 Young 72 7Document6 pagesHematology 2006 Young 72 7Indira DeviPas encore d'évaluation
- ZH 800906003455Document8 pagesZH 800906003455Dung TranPas encore d'évaluation
- 2 Haematological Profile in Children With Protein PDFDocument7 pages2 Haematological Profile in Children With Protein PDFAma BeluPas encore d'évaluation
- FA_Metabolomic and biochemical characterization of a new model of the transition of acute kidney injury to chronic kidney disease induced by folic acidDocument26 pagesFA_Metabolomic and biochemical characterization of a new model of the transition of acute kidney injury to chronic kidney disease induced by folic acidCher IshPas encore d'évaluation
- Chemical Constituents and Biological Activity of Medicinal Plants Used For The Management of Sickle Cell Disease - A ReviewDocument25 pagesChemical Constituents and Biological Activity of Medicinal Plants Used For The Management of Sickle Cell Disease - A ReviewIrfan Ullah WaniPas encore d'évaluation
- Pediatric Systemic Lupus Erythematosus Associated With AutoimmuneDocument9 pagesPediatric Systemic Lupus Erythematosus Associated With AutoimmunegistaluvikaPas encore d'évaluation
- Orantes Navarro 2017Document6 pagesOrantes Navarro 2017Maria Clara Formolo SouzaPas encore d'évaluation
- Prevalence of Anemia Among Pregnant Women Attending Antenatal Care at Tikur Anbessa Specialized Hospital Addis Ababa Ethiopia 2329 8790.1000125Document6 pagesPrevalence of Anemia Among Pregnant Women Attending Antenatal Care at Tikur Anbessa Specialized Hospital Addis Ababa Ethiopia 2329 8790.1000125thaynara robertoPas encore d'évaluation
- Assessment of Membrane Erythrocyte Cholesterol Level in Sickle Cell DiseaseDocument9 pagesAssessment of Membrane Erythrocyte Cholesterol Level in Sickle Cell DiseaseSely wulan widya ningsihPas encore d'évaluation
- Protective Effects of Hydro Ethanolic Leaf Extract Based FormulationDocument10 pagesProtective Effects of Hydro Ethanolic Leaf Extract Based FormulationMariaJesusWongPas encore d'évaluation
- Dental Journal: Oral Lesions As A Clinical Sign of Systemic Lupus ErythematosusDocument6 pagesDental Journal: Oral Lesions As A Clinical Sign of Systemic Lupus ErythematosusIin Ardi APas encore d'évaluation
- Pone 0234306 PDFDocument18 pagesPone 0234306 PDFyenny handayani sihitePas encore d'évaluation
- An Update On Diabetes Mellitus: June 2018Document12 pagesAn Update On Diabetes Mellitus: June 2018Siti AnisaPas encore d'évaluation
- Acute Liver FailureDocument12 pagesAcute Liver FailureRodrigo BenitezPas encore d'évaluation
- Diagnosing and predicting outcome in leptospirosis: the need for clinically relevant basic sciences researchDocument3 pagesDiagnosing and predicting outcome in leptospirosis: the need for clinically relevant basic sciences researchAnonymous fOcbAtPas encore d'évaluation
- Cirrosis Biliar PrimariaDocument10 pagesCirrosis Biliar PrimariaGerardo CorreaPas encore d'évaluation
- Sickle Cell Anemia A Succinct Look Into Recently Tested Disease Modifying InterventionsDocument10 pagesSickle Cell Anemia A Succinct Look Into Recently Tested Disease Modifying InterventionsAthenaeum Scientific PublishersPas encore d'évaluation
- Wilson Update FFF BunDocument20 pagesWilson Update FFF BunAlin CiubotaruPas encore d'évaluation
- English For Medical Ii Plenary I: "Aplastic Anemia"Document13 pagesEnglish For Medical Ii Plenary I: "Aplastic Anemia"Kang Wisit TheaPas encore d'évaluation
- Acute Kidney Injury in COVID-19: Emerging Evidence of A Distinct PathophysiologyDocument4 pagesAcute Kidney Injury in COVID-19: Emerging Evidence of A Distinct PathophysiologyxtinePas encore d'évaluation
- Amyloid Beta Multiple Mechanisms of Toxicity and Only Some Protective EffectsDocument16 pagesAmyloid Beta Multiple Mechanisms of Toxicity and Only Some Protective EffectsEvernim OmpacanPas encore d'évaluation
- Lola SeminarDocument12 pagesLola SeminarSOMOSCOPas encore d'évaluation
- AmebiasisDocument10 pagesAmebiasisMuhammad RifkiPas encore d'évaluation
- Microglial Inflammation and Phagocytosis in Alzheimer's Disease: Potential Therapeutic TargetsDocument18 pagesMicroglial Inflammation and Phagocytosis in Alzheimer's Disease: Potential Therapeutic TargetsChristina MountakiPas encore d'évaluation
- Immunomodulatory Therapy For The Management of Loiasis PDFDocument5 pagesImmunomodulatory Therapy For The Management of Loiasis PDFLucchéri Ndong AkomezoghePas encore d'évaluation
- 1768 PDFDocument2 pages1768 PDFAndhiky Raymonanda MadangsaiPas encore d'évaluation
- Penugasan - Diagnosis and Treatment of Aplastic AnemiaDocument22 pagesPenugasan - Diagnosis and Treatment of Aplastic AnemiaokerPas encore d'évaluation
- Nefrotik SyndDocument13 pagesNefrotik SyndJorianditha RamadhanPas encore d'évaluation
- Mas SleDocument12 pagesMas SlerahPas encore d'évaluation
- FEBS Letters - 2013 - Joshi - High Glucose Modulates IL 6 Mediated Immune Homeostasis Through Impeding NeutrophilDocument6 pagesFEBS Letters - 2013 - Joshi - High Glucose Modulates IL 6 Mediated Immune Homeostasis Through Impeding NeutrophilFlávia PampolhaPas encore d'évaluation
- Percário_etal_Oxidative Stress in MalariaDocument27 pagesPercário_etal_Oxidative Stress in MalariaNathália AraújoPas encore d'évaluation
- Anemia in Cancer Some Pathophysiological AspectsDocument6 pagesAnemia in Cancer Some Pathophysiological AspectsCindyGaniPas encore d'évaluation
- Article 1576845020 PDFDocument5 pagesArticle 1576845020 PDFFatima Habib JajjaPas encore d'évaluation
- NOMA ArticuloDocument3 pagesNOMA ArticuloERICAPas encore d'évaluation
- The Relationship Between Anaemia and Malaria: Apparently Simple, Yet ControversialDocument3 pagesThe Relationship Between Anaemia and Malaria: Apparently Simple, Yet ControversialAuliangi TamayoPas encore d'évaluation
- NIH Public Access: Author ManuscriptDocument15 pagesNIH Public Access: Author ManuscriptAdichita Khaira RidatasatriaPas encore d'évaluation
- Kaposi SarcomaDocument14 pagesKaposi Sarcomaedgar mandengPas encore d'évaluation
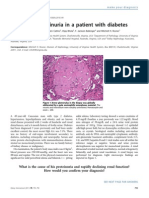
- The Case - Proteinuria in A Patient With DiabetesDocument2 pagesThe Case - Proteinuria in A Patient With Diabetesdrgawad2797Pas encore d'évaluation
- Pharmaceutical Sciences: A Case-Control Study of Prevalence of Anaemia Among Patients With Type 2 DiabetesDocument5 pagesPharmaceutical Sciences: A Case-Control Study of Prevalence of Anaemia Among Patients With Type 2 DiabetesiajpsPas encore d'évaluation
- Impact of Malaria Parasitemia On Some Liver Enzymes Among Adults Patients Attending University of Maiduguri Teaching Hospital, Maiduguri, Borno State-NigeriaDocument8 pagesImpact of Malaria Parasitemia On Some Liver Enzymes Among Adults Patients Attending University of Maiduguri Teaching Hospital, Maiduguri, Borno State-NigeriaAcademic JournalPas encore d'évaluation
- The Aetiology and Management of Atrophic RhinitisDocument11 pagesThe Aetiology and Management of Atrophic Rhinitisanoop_aiims1Pas encore d'évaluation
- FA__Folic acid-induced animal model of kidney diseaseDocument14 pagesFA__Folic acid-induced animal model of kidney diseaseCher IshPas encore d'évaluation
- Editorial: DNA Damage: Health and LongevityDocument2 pagesEditorial: DNA Damage: Health and LongevityAngelica CobosPas encore d'évaluation
- The Silent Anaemia Epidemic in ChildrenDocument5 pagesThe Silent Anaemia Epidemic in ChildrenInternational Journal of Innovative Science and Research TechnologyPas encore d'évaluation
- Loss-Of-Function Mutations in CARD14 Are Associated With A Severe Variant of Atopic DermatitisDocument20 pagesLoss-Of-Function Mutations in CARD14 Are Associated With A Severe Variant of Atopic DermatitisneleaPas encore d'évaluation
- TKF 2Document4 pagesTKF 2api-257045584Pas encore d'évaluation
- Diagnosis and Management of Autoimmune Hepatitis: A Clinical GuideD'EverandDiagnosis and Management of Autoimmune Hepatitis: A Clinical GuideMark W. RussoPas encore d'évaluation
- Circulationaha 110 009449 PDFDocument12 pagesCirculationaha 110 009449 PDFAhmad ThotuchingPas encore d'évaluation
- European Heart Journal Provides Guidance on Diagnosis and Management of Deep Vein ThrombosisDocument13 pagesEuropean Heart Journal Provides Guidance on Diagnosis and Management of Deep Vein ThrombosisDinPas encore d'évaluation
- Management of Spontaneous Reperfusion and Late Arrival - Héctor Bueno, MD PDFDocument19 pagesManagement of Spontaneous Reperfusion and Late Arrival - Héctor Bueno, MD PDFAhmad ThotuchingPas encore d'évaluation
- Morning Report: 82-Year-Old Man With Chest PainDocument22 pagesMorning Report: 82-Year-Old Man With Chest PainAhmad ThotuchingPas encore d'évaluation
- Group 8 A2 Modules 5 7 IMMUNOHEMATOLOGYDocument4 pagesGroup 8 A2 Modules 5 7 IMMUNOHEMATOLOGYKrystelle Anne PenaflorPas encore d'évaluation
- Relationship of Cell-Free Hemoglobin To Impaired Endothelial Nitric Oxide Bioavailability and Perfusion in Severe Falciparum MalariaDocument8 pagesRelationship of Cell-Free Hemoglobin To Impaired Endothelial Nitric Oxide Bioavailability and Perfusion in Severe Falciparum MalariaAhmad ThotuchingPas encore d'évaluation
- Proteins in Serum & UrineDocument66 pagesProteins in Serum & Urinekiedd_04100% (1)
- 9.1 Liver Function TestsDocument8 pages9.1 Liver Function TestsGio Joaquin SantosPas encore d'évaluation
- Congenital and Aquired Hemolytic Anemias QuestionsDocument36 pagesCongenital and Aquired Hemolytic Anemias QuestionstoniassafPas encore d'évaluation
- Ninja Nerd AnemiaDocument13 pagesNinja Nerd AnemiaAndra Bauer100% (3)
- Plasma Proteins 1Document46 pagesPlasma Proteins 1Faisal_Khatib_juPas encore d'évaluation
- Hemolytic AnemiaDocument7 pagesHemolytic AnemiaVictoria BrandedBagPas encore d'évaluation
- Anemias ChartDocument14 pagesAnemias ChartM Patel100% (2)
- ASCP Questions (Autosaved) - 1Document56 pagesASCP Questions (Autosaved) - 1Glenn Perez100% (16)
- MLAB 2401: Clinical Chemistry Keri Brophy-Martinez: Plasma ProteinsDocument19 pagesMLAB 2401: Clinical Chemistry Keri Brophy-Martinez: Plasma ProteinsLoulou ApolloPas encore d'évaluation
- Hematology Mudit Khanna PDFDocument40 pagesHematology Mudit Khanna PDFSylvia DiamondPas encore d'évaluation
- Rodak's Hematology Clinical Principles and Applications Ch 12 Case StudyDocument25 pagesRodak's Hematology Clinical Principles and Applications Ch 12 Case StudyBernard Garcia OledanPas encore d'évaluation
- 2.2. Biosintesis Hemoglobin PDFDocument48 pages2.2. Biosintesis Hemoglobin PDFIkaTriRahayuPas encore d'évaluation
- Preanalytical ErrorsDocument99 pagesPreanalytical ErrorsSulakshmi KurlekarPas encore d'évaluation
- Red and White Blood Cells TurnoverDocument4 pagesRed and White Blood Cells TurnoverNabillaMerdikaPutriKusumaPas encore d'évaluation
- Anemia: Differential DiagnosisDocument7 pagesAnemia: Differential DiagnosisZaman MigozaradPas encore d'évaluation
- 003assessment Exam ImmunohemaDocument41 pages003assessment Exam ImmunohemaFrankenstein MelancholyPas encore d'évaluation
- Saint Luke's College Of: NursingDocument5 pagesSaint Luke's College Of: NursingApril Mae Magos LabradorPas encore d'évaluation
- Rodak's Hematology 6th Edition Test BankDocument255 pagesRodak's Hematology 6th Edition Test Bankromerolourdes4Pas encore d'évaluation
- Hemoglobin Structure and Function in 40 CharactersDocument4 pagesHemoglobin Structure and Function in 40 Charactersrona hilarioPas encore d'évaluation
- ABP3 IntoxmetalespesadosDocument18 pagesABP3 IntoxmetalespesadosFernandoLuyoPas encore d'évaluation
- Hematology Part 1 NotesDocument23 pagesHematology Part 1 NotesDr. Benson BenjaminPas encore d'évaluation
- RBC, Structure and FunctionDocument17 pagesRBC, Structure and FunctionAsyha KantifaPas encore d'évaluation
- Microcytic AnemiaDocument6 pagesMicrocytic AnemiaharideepPas encore d'évaluation
- Anemia Hemolitica Congenita y Adq Ped in ReviewDocument14 pagesAnemia Hemolitica Congenita y Adq Ped in ReviewEliana LopezPas encore d'évaluation
- Cc1 Quiz Plasma Proteins Blank 1 and 2Document2 pagesCc1 Quiz Plasma Proteins Blank 1 and 2Treyton Sekani LopezPas encore d'évaluation
- September - 6 - 2021-For - Students (HEMA)Document24 pagesSeptember - 6 - 2021-For - Students (HEMA)Christian John V. CamorahanPas encore d'évaluation
- Pathology - Chapter 14Document14 pagesPathology - Chapter 14Cory GrayPas encore d'évaluation
- 3f - 1 (Division of Task)Document33 pages3f - 1 (Division of Task)Trisha ArtosPas encore d'évaluation