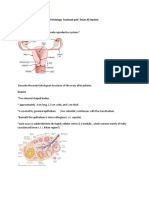
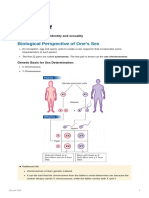
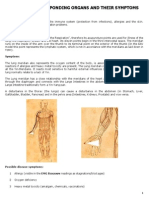
Vous aimerez peut-être aussi
- Williams 03 - Implantation, Embryogenesis, Placental DevelopmentDocument14 pagesWilliams 03 - Implantation, Embryogenesis, Placental DevelopmentAhmed Abou SeifPas encore d'évaluation
- Implantataion Placental Development and AbnormalitiesDocument62 pagesImplantataion Placental Development and AbnormalitiesIsabel CastilloPas encore d'évaluation
- Organa Genitalia Feminina: Oleh Dr. Hendra SutardhioDocument90 pagesOrgana Genitalia Feminina: Oleh Dr. Hendra SutardhioMirah WilayadiPas encore d'évaluation
- CH 5 Implantation, Placental Dev (Part1)Document36 pagesCH 5 Implantation, Placental Dev (Part1)Tengku Chairannisa PutriPas encore d'évaluation
- Organa Genitalia Feminina: Oleh Dr. Hendra SutardhioDocument90 pagesOrgana Genitalia Feminina: Oleh Dr. Hendra SutardhioNathasya PakpahanPas encore d'évaluation
- Maternal Placental Foetal UnitDocument10 pagesMaternal Placental Foetal UnitFestus SinoPas encore d'évaluation
- Chapt17 Placenta PDFDocument12 pagesChapt17 Placenta PDFAnonymous 7resQ9Pas encore d'évaluation
- Implantation and PlacentaDocument52 pagesImplantation and PlacentaMs BadooPas encore d'évaluation
- 1.1c Implantation & Placental DevelopmentDocument5 pages1.1c Implantation & Placental DevelopmentBea Samonte100% (1)
- Imported FileDocument18 pagesImported Fileg8pn6pt8nhPas encore d'évaluation
- Obstetrics: The Normal Menstrual CycleDocument4 pagesObstetrics: The Normal Menstrual Cycleapi-3829364Pas encore d'évaluation
- Abnormal ImplantationDocument3 pagesAbnormal ImplantationMuhammet Fatih CantepePas encore d'évaluation
- Implantasi Dan Perkembangan PlasentaDocument18 pagesImplantasi Dan Perkembangan PlasentaHei KamuPas encore d'évaluation
- Histo FemDocument9 pagesHisto FemzaidPas encore d'évaluation
- 1 - Physiology of The Menstrual CycleDocument17 pages1 - Physiology of The Menstrual CycleMalak Ahmed 2295Pas encore d'évaluation
- Capacitation: Acrosome ReactionDocument13 pagesCapacitation: Acrosome Reaction尤淳敏Pas encore d'évaluation
- Naim LBM 1-1Document38 pagesNaim LBM 1-1dimasPas encore d'évaluation
- Mechanism of The Menstrual CycleDocument4 pagesMechanism of The Menstrual Cyclemartin magdenoskiPas encore d'évaluation
- Anatomy and Physiology IIDocument129 pagesAnatomy and Physiology IIkenmanikesePas encore d'évaluation
- Urogenital Development & Associated AnomaliesDocument114 pagesUrogenital Development & Associated AnomaliesvictorPas encore d'évaluation
- 5 Maternal Adaptations To Pregnancy by Dr. Catherine Torres-JisonDocument75 pages5 Maternal Adaptations To Pregnancy by Dr. Catherine Torres-JisonKevin AgbonesPas encore d'évaluation
- Chapter 29 - Development and Inheritance: ST ND RD STDocument5 pagesChapter 29 - Development and Inheritance: ST ND RD STtomorrow.today.yesterday .yesterdayPas encore d'évaluation
- Pathology of Preeclampsia (Autosaved) (Autosaved)Document59 pagesPathology of Preeclampsia (Autosaved) (Autosaved)ck4realPas encore d'évaluation
- Female Reproductive System: UterusDocument44 pagesFemale Reproductive System: UterusBeni KurniawanPas encore d'évaluation
- 1st, 2nd & 3rdweeksDocument9 pages1st, 2nd & 3rdweeksPristine KrPas encore d'évaluation
- Williams Chapters 3 & 4Document2 pagesWilliams Chapters 3 & 4DrÄlamuri ßashaPas encore d'évaluation
- 009 - Reproductive HistologyDocument10 pages009 - Reproductive HistologycharlieholdenPas encore d'évaluation
- Endometrium by Shanojan ThiyagalingamDocument3 pagesEndometrium by Shanojan ThiyagalingamDr. Shanojan Thiyagalingam, FRCPC FACPPas encore d'évaluation
- EMBRYOLOGYDocument30 pagesEMBRYOLOGYMSc. PreviousPas encore d'évaluation
- Fertilization and ImplantationDocument3 pagesFertilization and ImplantationTandingco, Olivia Mari H.Pas encore d'évaluation
- Perkembangan Plasenta (Autosaved) .En - IdDocument18 pagesPerkembangan Plasenta (Autosaved) .En - IdFATIMAHPas encore d'évaluation
- Menstrual Cycle: Definition of MenstruationDocument5 pagesMenstrual Cycle: Definition of MenstruationMuhammad Azam Mohd NizaPas encore d'évaluation
- Female Reproductive SystemDocument15 pagesFemale Reproductive Systemada.codreanuPas encore d'évaluation
- Fertilization and Implantation: Dr. Madhan KumarDocument47 pagesFertilization and Implantation: Dr. Madhan KumarTan DesmondPas encore d'évaluation
- Obstetricia 2019Document150 pagesObstetricia 2019Sara BalderasPas encore d'évaluation
- The Three Villus TypesDocument12 pagesThe Three Villus TypesMarhina Asarabi MukimPas encore d'évaluation
- Menstrual Cycle and Implantation)Document19 pagesMenstrual Cycle and Implantation)Ragdha MehdawiPas encore d'évaluation
- Female Genital Tract Basic HistologyDocument40 pagesFemale Genital Tract Basic HistologySantoshini RaviPas encore d'évaluation
- Fetal and Placental Development - FETAL Sac, Umbilical Cord .Abnormalities of PlacentaDocument150 pagesFetal and Placental Development - FETAL Sac, Umbilical Cord .Abnormalities of PlacentaKripa SusanPas encore d'évaluation
- Placental Development - Implantation - Transfer - Clinical RelevanceDocument3 pagesPlacental Development - Implantation - Transfer - Clinical RelevanceLiridon SopajPas encore d'évaluation
- Development of The Placenta and Its FunctionDocument27 pagesDevelopment of The Placenta and Its FunctionColince johnsonPas encore d'évaluation
- Female Genital System PDFDocument61 pagesFemale Genital System PDFWaleed AbdelallPas encore d'évaluation
- Fertilization and Blastocyst FormationDocument1 pageFertilization and Blastocyst FormationTyler KingPas encore d'évaluation
- Female Reproductive System Histology LectureDocument46 pagesFemale Reproductive System Histology LectureMwaePas encore d'évaluation
- Menstrual Cycle Dr. Hani MahdiDocument80 pagesMenstrual Cycle Dr. Hani MahdiArwa Qishta100% (1)
- Good Afternoon: Prepared byDocument102 pagesGood Afternoon: Prepared byKrishna PatelPas encore d'évaluation
- Lecture - 2 - 3 - Goal - Fertilization - Placentation - LactationDocument4 pagesLecture - 2 - 3 - Goal - Fertilization - Placentation - LactationAPOORVA SHANDILYAPas encore d'évaluation
- Parts of The SystemDocument14 pagesParts of The SystembookonscribdPas encore d'évaluation
- Conception and Diagnosis of Early PregnancyDocument48 pagesConception and Diagnosis of Early PregnancyjerrydanfordfxPas encore d'évaluation
- Placenta and Fetal Membranes 2010 1Document22 pagesPlacenta and Fetal Membranes 2010 1Qaiser InayatPas encore d'évaluation
- 1-4 Fertilization & Fetal DevelopmentDocument9 pages1-4 Fertilization & Fetal DevelopmentElla DionedaPas encore d'évaluation
- Charles Z. Ariola JR., MSN., LPT, Rn. Instructor IDocument22 pagesCharles Z. Ariola JR., MSN., LPT, Rn. Instructor ICharlz ZipaganPas encore d'évaluation
- Prof. Dr. Çiğdem ElmasDocument49 pagesProf. Dr. Çiğdem ElmasMuhammet Fatih CantepePas encore d'évaluation
- Menstruation & OvulationDocument42 pagesMenstruation & Ovulationneilesh300Pas encore d'évaluation
- Fetal Membranes IMBBSDocument26 pagesFetal Membranes IMBBSSubham YadavPas encore d'évaluation
- Embryology Type WrittenDocument74 pagesEmbryology Type WrittenAnonymous QqyLDoW1Pas encore d'évaluation
- MCH 314 Part ADocument5 pagesMCH 314 Part AAnifowose samsonPas encore d'évaluation
- Fallopian Tube DisordersDocument20 pagesFallopian Tube DisordersAndreas HaryonoPas encore d'évaluation
- Fertilization: Glora P. de Leon, RN, RM, ManDocument103 pagesFertilization: Glora P. de Leon, RN, RM, ManJoan VillafrancaPas encore d'évaluation
- Normal Menstrual Cycle The ProcessDocument6 pagesNormal Menstrual Cycle The ProcessBrett StevensonPas encore d'évaluation
- The Endocrine System: Seventh EditionDocument40 pagesThe Endocrine System: Seventh EditionAngelaPas encore d'évaluation
- Complete Androgen Insensitivity Syndrome: Garry L.WarneDocument29 pagesComplete Androgen Insensitivity Syndrome: Garry L.WarnelabuenoPas encore d'évaluation
- Biological Perspective of One's SexDocument13 pagesBiological Perspective of One's SexBIANCA NICOLE DAGUDAGPas encore d'évaluation
- Bioidentical Hormone TherapyDocument8 pagesBioidentical Hormone TherapyKorry Meliana PangaribuanPas encore d'évaluation
- Endocrine Glands Mini ProjectDocument7 pagesEndocrine Glands Mini ProjectJoy-Ann NuptialPas encore d'évaluation
- Material On PHYSIOLOGICAL CHANGES DURING PREGNANCYDocument21 pagesMaterial On PHYSIOLOGICAL CHANGES DURING PREGNANCYAnuradha MauryaPas encore d'évaluation
- Sex HormonesDocument442 pagesSex HormonesMITHUN CHATTERJEEPas encore d'évaluation
- Sex Hormone Synthesis, Regulation, and Function - McMaster Pathophysiology ReviewDocument7 pagesSex Hormone Synthesis, Regulation, and Function - McMaster Pathophysiology ReviewWahyu W'cas PamungkasPas encore d'évaluation
- Oecd 407Document13 pagesOecd 407Susan EliasPas encore d'évaluation
- Greendale-1999-The MenopauseDocument10 pagesGreendale-1999-The MenopauseRya RatnaPas encore d'évaluation
- Phytoestrogen and Ellagic Acid Sherlyn GeveroDocument19 pagesPhytoestrogen and Ellagic Acid Sherlyn GeveroAllysa Mae LavarnezPas encore d'évaluation
- DPC-3 Word - 1Document42 pagesDPC-3 Word - 1charlienitin1325Pas encore d'évaluation
- PhysioEx Exercise 4 Activity 3Document3 pagesPhysioEx Exercise 4 Activity 3PabloAndresPalaciosAgilaPas encore d'évaluation
- An in Vitro Study On Antiproliferative and Antiestrogenic Effects ofDocument5 pagesAn in Vitro Study On Antiproliferative and Antiestrogenic Effects ofJose Alberto PbPas encore d'évaluation
- Breast Cancer A Review of Risk FactorsDocument7 pagesBreast Cancer A Review of Risk FactorsKIU PUBLICATION AND EXTENSIONPas encore d'évaluation
- Pharmacognosy-Large FontDocument35 pagesPharmacognosy-Large FontArantxa HilarioPas encore d'évaluation
- Gynecology PDFDocument85 pagesGynecology PDFrachaPas encore d'évaluation
- Aqa Combisci Bio Paper 2 1650625 PDFDocument6 pagesAqa Combisci Bio Paper 2 1650625 PDFCuthbert YedwaPas encore d'évaluation
- 20 Bodybuilding SecretsDocument42 pages20 Bodybuilding SecretsMalal QuechuaPas encore d'évaluation
- Williams Ob Overall ReviewerDocument128 pagesWilliams Ob Overall ReviewerCamilla FernandezPas encore d'évaluation
- The Climacteric Lecture 10Document9 pagesThe Climacteric Lecture 10Miss Mona100% (1)
- Steroids From Physiology To Clinical MedicineDocument220 pagesSteroids From Physiology To Clinical MedicineRenan FernandesPas encore d'évaluation
- What Is Desoxy DHEA and How Does It WorkDocument2 pagesWhat Is Desoxy DHEA and How Does It WorkdenovonutritionPas encore d'évaluation
- Meridians Organs SymptomsDocument23 pagesMeridians Organs SymptomsFlorina Stefan100% (3)
- New Developments in The Medical & Surgical Therapy For FibroidsDocument20 pagesNew Developments in The Medical & Surgical Therapy For FibroidsernestosandPas encore d'évaluation
- Telescopium DewasaDocument11 pagesTelescopium DewasaBayu KrisnaPas encore d'évaluation
- Factors Affecting Bone GrowthDocument7 pagesFactors Affecting Bone GrowthNurafniPas encore d'évaluation
- Sarah Richardson - Plasticity and Programming (2017)Document24 pagesSarah Richardson - Plasticity and Programming (2017)SofíaRBPas encore d'évaluation
- GPDocument311 pagesGPNAVEENPas encore d'évaluation