Vous aimerez peut-être aussi
- Criterio Diagnostico Tipos de EsquizofreniaDocument3 pagesCriterio Diagnostico Tipos de EsquizofreniastevemburgPas encore d'évaluation
- Trastorno de EsquizofreniaDocument3 pagesTrastorno de EsquizofreniastevemburgPas encore d'évaluation
- Psicofarmacos PsiquiatriaDocument94 pagesPsicofarmacos Psiquiatriastevemburg100% (1)
- Actitudes Avanzadas de AVCADocument30 pagesActitudes Avanzadas de AVCAstevemburgPas encore d'évaluation
- Orina TrabajoDocument9 pagesOrina TrabajostevemburgPas encore d'évaluation
- Inhibidores de La Síntesis y Liberación de ProstaglandinaDocument6 pagesInhibidores de La Síntesis y Liberación de ProstaglandinastevemburgPas encore d'évaluation
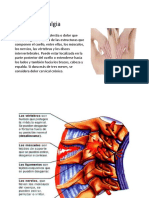
- CervicalgiaDocument25 pagesCervicalgiastevemburg100% (1)
- Formato Apa Anemia FerroenicaDocument20 pagesFormato Apa Anemia Ferroenicastevemburg100% (1)
- Inmunidad Respuesta Inmunitaria Frente A HongosDocument6 pagesInmunidad Respuesta Inmunitaria Frente A HongosAngelo Di RienzoPas encore d'évaluation
- PC3 Practica y TeoriaDocument113 pagesPC3 Practica y TeoriaXimena GarciaPas encore d'évaluation
- Preguntas MHCDocument1 pagePreguntas MHCEdgar LopezPas encore d'évaluation
- Complejo Mayor de Histocompatibilidad y DesarrolloDocument12 pagesComplejo Mayor de Histocompatibilidad y DesarrolloJOSE DUBERLEE TAVARA VEGAPas encore d'évaluation
- CAPITULO 6 Enfermedades de La InmunidadDocument24 pagesCAPITULO 6 Enfermedades de La InmunidadLuis PortuguezPas encore d'évaluation
- Presentacion AntigenicaDocument21 pagesPresentacion AntigenicaMassiel AguilarPas encore d'évaluation
- Invasiones BacterianasDocument10 pagesInvasiones BacterianasJose Antonio VenacostaPas encore d'évaluation
- Mecanismos de Evasión Inmunitaria de La LeishmaniasisDocument7 pagesMecanismos de Evasión Inmunitaria de La LeishmaniasisIsaPas encore d'évaluation
- Artículo Complejo Mayor de HistocompatibilidadDocument4 pagesArtículo Complejo Mayor de HistocompatibilidadRebeca Saltos CandelaPas encore d'évaluation
- MHC en El Rechazo de TrasplantesDocument3 pagesMHC en El Rechazo de TrasplantesmishellPas encore d'évaluation
- Fisiologia y Fisica BiológicaDocument454 pagesFisiologia y Fisica BiológicaGuilherme de Souza Peixoto DuartePas encore d'évaluation
- InmunologiaDocument43 pagesInmunologiatitoxxxyyy100% (1)
- Fundamentos Sistema InmunitarioDocument23 pagesFundamentos Sistema InmunitarioLaura Daniela Sierra GuevaraPas encore d'évaluation
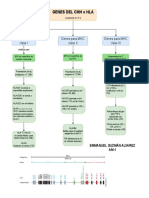
- Genes de HLA o MHCDocument1 pageGenes de HLA o MHCEmmanuel GPas encore d'évaluation
- p1.3 VNR Sistema Inmune 13-03-18 Dra Celeste RodriguezDocument8 pagesp1.3 VNR Sistema Inmune 13-03-18 Dra Celeste RodriguezRodrigo Ancalle ZuñigaPas encore d'évaluation
- Departamental Inmuno IIIDocument8 pagesDepartamental Inmuno IIIGus TorresPas encore d'évaluation
- Complejo Principal de HistocompatibilidadDocument45 pagesComplejo Principal de HistocompatibilidadDánae Jazmín Llanca BravoPas encore d'évaluation
- 210 PreguntasDocument26 pages210 PreguntaseddumedinaPas encore d'évaluation
- Cuestionario 1 - Técnicas de InmunologiaDocument18 pagesCuestionario 1 - Técnicas de InmunologiaJocelyn OrellanaPas encore d'évaluation
- Clase#5clasificacion de La Diabetes MellitusDocument32 pagesClase#5clasificacion de La Diabetes MellitusAljavainPas encore d'évaluation
- SALAZAR CORTES DOMENICA ELIZABETH Haplotipo de HLADocument4 pagesSALAZAR CORTES DOMENICA ELIZABETH Haplotipo de HLAMARTIN ALEJANDRO SALAZAR CORTESPas encore d'évaluation
- Semana 10Document45 pagesSemana 10Kaylee Lopez LinguinniPas encore d'évaluation
- Resumen Cap 6 InmunoDocument6 pagesResumen Cap 6 Inmunobopeepout papuPas encore d'évaluation
- CAPÍTULO 8 - Desarrollo de Linfocitos TDocument40 pagesCAPÍTULO 8 - Desarrollo de Linfocitos TAlejandra MincholaPas encore d'évaluation
- Inmunologia pc4Document158 pagesInmunologia pc4Victoria Zuñiga EscatePas encore d'évaluation
- 4-Procesamiento de AntígenosDocument22 pages4-Procesamiento de AntígenosJOSE DUBERLEE TAVARA VEGAPas encore d'évaluation
- Activación de Linfocitos T AlorreactivosDocument9 pagesActivación de Linfocitos T AlorreactivosManuel Rojas GonzaalesPas encore d'évaluation
- 9.3. Natural Killer Cells (Células Natural Killer)Document1 page9.3. Natural Killer Cells (Células Natural Killer)Marco Antonio LeónPas encore d'évaluation
- Inmunología MédicaDocument106 pagesInmunología MédicaCristóbal Muñoz PérezPas encore d'évaluation
- Clase Caracteristicas Generales de Las Barreras Defensivas Del Cuerpo HumanoDocument77 pagesClase Caracteristicas Generales de Las Barreras Defensivas Del Cuerpo HumanoKarina DonosoPas encore d'évaluation