Académique Documents
Professionnel Documents
Culture Documents
Neurociencia Aplicada 2007 PDF
Transféré par
Brayan CorderoTitre original
Copyright
Formats disponibles
Partager ce document
Partager ou intégrer le document
Avez-vous trouvé ce document utile ?
Ce contenu est-il inapproprié ?
Signaler ce documentDroits d'auteur :
Formats disponibles
Neurociencia Aplicada 2007 PDF
Transféré par
Brayan CorderoDroits d'auteur :
Formats disponibles
Parte I
Los componentes del sistema nervioso
y su comunicación
1 Biología de las células nerviosas
2 Generación y conducción de potenciales en el sistema nervioso
3 Transmisión sináptica
Neurociencia aplicada ©2007. Editorial Médica Panamericana
1
Biología de las células nerviosas
En las neuronas existen regiones
funcionalmente diferenciadas
En una neurona típica pueden identificarse mor-
fológicamente cuatro regiones: a) el cuerpo celu-
lar, llamado también soma o pericarion, b) las
dendritas, c) el axón y d) las terminales axónicas o
sinápticas (fig. 1-1).
La función principal de las neuronas es la genera-
ción de señales eléctricas, y en esta actividad cada una
de las partes señaladas tiene un papel específico. Fig. 1-1. Neurona típica con las sinapsis que recibe. De iz-
El cuerpo celular (o pericarion) constituye el centro quierda a derecha, axodendrítica, axosomática, axoaxónica
funcional y metabólico de la neurona y contiene tres proximal y axoaxónica distal. Esta última en general inhibito-
organelas fundamentales: ria, con participación en la inhibición presináptica.
• El núcleo celular, que en las neuronas, a diferencia
de otras células, es de gran tamaño. tamaño de una pelota de tenis, el axón tendría unos 2
• El retículo endoplasmático, donde se sintetizan las km de longitud y el árbol dendrítico ocuparía el volu-
proteínas de membrana y secretorias. men de una habitación de unos 4 × 4 metros. Esto des-
• El aparato de Golgi, donde se realiza el procesado taca la arbitrariedad de esquemas neuronales como los
de los componentes de membrana y secretorios. de la figura 1-1: el árbol dendrítico es de una extraor-
dinaria importancia para la neurona, no reflejado en
Las dendritas son arborizaciones del cuerpo celular los esquemas habituales.
que desempeñan el papel de zona receptora principal Cuando los axones son gruesos están rodeados de
para la neurona. una vaina aislante, la mielina, provista por las células
El axón, proceso tubular que puede alcanzar distan- de Schwann en la periferia y por la oligodendroglia en
cias considerables, actúa como la unidad conductiva el SNC. La vaina de mielina es esencial para la con-
de la neurona. ducción de alta velocidad y se halla interrumpida en
Los tamaños relativos del cuerpo neuronal, de las los nervios periféricos, a intervalos regulares, por los
dendritas y del axón son variables de neurona a neuro- nodos de Ranvier.
na. En muchos casos, el axón puede superar en varios Las terminales axónicas o sinápticas constituyen
órdenes de magnitud el diámetro del cuerpo celular. los elementos de transmisión de la neurona. A través
Como caso extremo puede mencionarse el de una mo- de ellas, una neurona contacta y transmite información
toneurona lumbar que inerve algún músculo del pie. Si a la zona receptiva de otra neurona, o de una célula
se ampliara el cuerpo celular de esta motoneurona al efectora (p. ej., muscular).
Neurociencia aplicada ©2007. Editorial Médica Panamericana
4 Parte I - Los componentes del sistema nervioso y su comunicación
La zona de contacto se llama sinapsis. Cuando se nas cerebrales, mientras que en las neuronas monoa-
trata de una neurona, la zona postsináptica se ubica en minérgicas presentan una profusa arborización “en
las dendritas y, con menos frecuencia, en el cuerpo telaraña”, que conecta con numerosas áreas cerebra-
neuronal o en las porciones iniciales o finales del les muchas veces alejadas entre sí.
axón. • Neuronas de axón corto, o de tipo Golgi II, que
En promedio, existen unos 1015 contactos sinápticos cumplen la función de interneuronas en circuitos lo-
en el cerebro humano adulto (es decir, unas 10.000 cales.
terminaciones sinápticas por neurona, aunque el nú-
mero de estas terminaciones varía notablemente de un Podemos así enunciar una regla elemental de forma-
tipo neuronal a otro). ción de los circuitos neuronales en el SNC: “dos neu-
Sobre la base del número de procesos originados en ronas × tres circuitos”.
el cuerpo neuronal, las neuronas se clasifican en tres
grupos:
Es decir, dos tipos neuronales (Golgi I y Golgi II)
generan los tres circuitos básicos:
• Unipolares.
• Bipolares.
• Multipolares. • Circuitos locales, formado por interneuronas.
• Circuitos de proyección o “punto a punto”, que
Las neuronas unipolares se encuentran en inverte- conectan circuitos locales lejanos entre sí.
brados y presentan un único proceso que da origen a • Circuitos “en telaraña”, que dan la base para
varias ramas. Estas ramas desempeñan la función de que modificaciones locales y aisladas se trans-
axón o de dendritas. En los mamíferos, la neurona sen- formen en estados globales del SNC, por ejem-
sorial primaria de los ganglios de las raíces dorsales es plo, la vigilia, el sueño lento y el sueño REM
una variante de la neurona unipolar, llamada seudou- (de rapid eye movements, movimientos oculares
nipolar (fig. 1-2), porque da origen a dos ramas fun- rápidos).
cionales, una periférica o dendrítica, y otra central que
constituye las raíces dorsales de los nervios espinales. Las células de la glía son el componente
Las neuronas bipolares son de soma ovoide con dos celular más abundante del SNC
procesos: periférico (de función dendrítica) y central
(o axonal). Las neuronas bipolares de la retina son un
ejemplo de esta clase de neuronas (véase fig. 1-2). El tipo celular más abundante en el SNC es el de las
Las neuronas multipolares son el tipo predominan- células de la glía, cuyo número excede unas 10-50 ve-
te en el SNC de los mamíferos. Presentan arborizacio- ces el de las neuronas. En general, las células gliales
nes dendríticas y, en general, un solo axón; las arbori- carecen de la propiedad de generar activamente seña-
zaciones dendríticas pueden emerger en todas las les eléctricas.
direcciones del cuerpo axonal. Son ejemplos de neu- Las células gliales tienen:
ronas multipolares las células piramidales de la corte-
za cerebral, las motoneuronas espinales y las células • Una función de soporte para las neuronas, semejan-
de Purkinje del cerebelo (figs. 1-2 y 1-3). te al papel del tejido conectivo en otros órganos.
De acuerdo con la longitud del axón, indicativa de la • La función de eliminación de productos de desecho
función que desempeñan, se distinguen dos tipos de del metabolismo neuronal, o de restos celulares lue-
neuronas: go de la lesión o muerte celular.
• La provisión de vaina de mielina (figs. 1-3 y 1-5).
• Neuronas de axón largo, o de tipo Golgi I, que • Una función de buffer espacial de K+ (fig. 1-4).
participan en la transferencia de información entre • Una función de guía para la migración neuronal
regiones cerebrales (p. ej., neuronas piramidales de durante el desarrollo.
proyección de la corteza cerebral), o que proveen un • Una función de nutrición neuronal, con la provi-
tono basal de excitación a amplias áreas cerebrales sión entre otros de lactato y glucosa (fig. 1-6).
(p. ej., neuronas monoaminérgicas “en telaraña” del • Función de captación de neurotransmisores (p. ej.,
tronco encefálico). La diferencia entre estos dos glutamato; fig. 1-6).
subgrupos de neuronas Golgi I es el grado de rami- • Una función activa de generación de señales de tipo
ficación del axón. En las neuronas de proyección, paracrino, como distintas citocinas. Este aspecto es
las ramificaciones se limitan a una o unas pocas zo- de vital importancia para entender los cuadros emo-
Neurociencia aplicada ©2007. Editorial Médica Panamericana
Biología de las células nerviosas 5
Fig. 1-2. Tipos de neuronas en distintas áreas del sistema nervioso central.
cionales que acompañan a las infecciones o al desa-
rrollo de tumores. La manera en que la reacción in-
mune periférica afecta al SNC es por acción de las
citocinas circulantes sobre células gliales a través de
los órganos circunventriculares.
• Una muy reciente función identificada para las célu-
las gliales es la de su capacidad de regeneración
neuronal. Este aspecto está siendo muy estudiado
y se inserta en la verificación de la capacidad del
SNC para reestablecer el stock neuronal de áreas
afectadas.
Las células gliales se dividen en los siguientes gru-
pos: a) macroglia, que comprende a los astrocitos, los
oligodendrocitos, las células de Schwann y los epen-
dimocitos. Es de origen ectodérmico, b) microglia,
que comprende fagocitos, que son parte del sistema
inmune. Es de origen mesodérmico.
Los astrocitos median las funciones gliales mencio-
nadas, salvo la de producir mielina, que es función Fig. 1-3. Pasos en la mielinización de un axón por la célula de
de la oligodendroglia en el SNC y de la célula de Schwann.
Neurociencia aplicada ©2007. Editorial Médica Panamericana
6 Parte I - Los componentes del sistema nervioso y su comunicación
Fig. 1-4. Función de buffer espacial de K+ de las células gliales. El catión que se acumula por la actividad neural se difunde por la
extremada permeabilidad de la membrana del astrocito.
Schwann en la periferia (figs. 1-3 y 1-5). La síntesis antígenos en neuronas que no implique participación
de mielina por los oligodendrocitos está, sin embargo, de las células de la glía.
bajo la regulación indirecta de los astrocitos, a través Durante el proceso temprano de mielinización, las
de una interacción de tipo paracrino. células de Schwann expresan una glucoproteína
Aunque los oligodendrocitos y las células de Sch- (MAG, myelin-associated glycoprotein) (sólo una par-
wann están específicamente encargados de la produc- te minoritaria en la mielina madura), que se encuentra
ción de la vaina de mielina, difieren entre sí en varios concentrada en la adyacencia inmediata de la mem-
aspectos funcionales. Existen unas 400-500 células de brana axonal. El MAG pertenece a una superfamilia
Schwann para envolver el axón periférico de una neu- de inmunoglobulinas implicadas en el reconocimiento
rona sensorial primaria del nervio femoral (de unos celular; otros miembros son el antígeno mayor de his-
0,5 metros de longitud, con distancia internodo de tocompatibilidad, la Po, los antígenos de superficie de
Ranvier de alrededor de 1 mm). En cambio, la prolon- los linfocitos T y las moléculas de adhesión de células
gación central de esa misma neurona sensorial está neurales.
contenida, junto con otras semejantes, en un único oli- Una enfermedad neurológica, la esclerosis en pla-
godendrocito (fig. 1-5). ca, se caracteriza por el desarrollo de autoanticuer-
Otra diferencia es que los genes que participan en la pos contra proteínas de la mielina. La principal pro-
síntesis de mielina en la célula de Schwann son activa- teína en la mielina periférica madura es llamada
dos por la presencia de axones, mientras que los de los “Po” y atraviesa la membrana celular de la célula de
oligodendrocitos lo son por la presencia de astrocitos. Schwann. Esta proteína pertenece también a la su-
Debe destacarse que no hay reacción fisiológica ante perfamilia de proteínas de reconocimiento celular.
Neurociencia aplicada ©2007. Editorial Médica Panamericana
Biología de las células nerviosas 7
Su función es la de interaccionar con moléculas
semejantes en el proceso de compactación de la mie-
lina.
En la parte central de la mielina (que carece de Po)
predomina un proteolípido (50% de la proteína pre-
sente). El resto de las proteínas mielínicas, tanto en
la parte central como en la periférica de la mielina,
son las conocidas como “proteína mielínica básica”,
y derivan de un mismo gen. Se puede desarrollar una
encefalomielitis alérgica experimental en ratas con
la inyección de antígenos de mielina y cuya evolu-
ción tiene las características de la enfermedad cróni-
ca humana.
La actividad neuronal, con la consiguiente acu-
mulación de K+ en el espacio extracelular, produce
la despolarización de las células gliales. Al ser la
membrana celular de los astrocitos permeable en
forma casi exclusiva al K+, este catión es captado
con facilidad por los astrocitos, con lo que se impi-
de una acumulación que resultaría peligrosa para
la función neuronal (función de “buffer espacial de
K+”) (véase fig. 1-4).
Se ha verificado que la conductancia al K+ difiere
entre las distintas regiones del astrocito y es muy
elevada en el pie vascular. En forma proporcional a
la actividad neuronal, la concentración extracelular
de K+ puede variar entre 4 y 10 mM (lo normal es Fig. 1-5. La célula de Schwann envuelve el axón periférico de
2,5 mM), que produce vasodilatación importante una neurona sensorial primaria. La prolongación central de esa
(50% de aumento del diámetro vascular cuando se misma neurona sensorial está envuelta por un oligodendrocito.
alcanzan 10 mM de K+). Al servir los pies vascula-
res (podocitos) de los astrocitos como “buffer espa-
cial” para el K+, proveen un mecanismo efectivo de
autorregulación del flujo sanguíneo cerebral. Co- El líquido cefalorraquídeo constituye
mo los astrocitos están conectados entre sí a través
de uniones estrechas, se forma entre ellos un amplio
la aproximación más cercana al líquido
sincitio funcional, con posibilidad de perder en otra intersticial cerebral y está separado de la
región el K+ ganado en una región celular (véase fig. circulación sistémica por dos barreras
1-4).
En los últimos 15 años se ha identificado toda la Además de la masa cerebral (unos 1.400 gra-
gama de canales dependientes del voltaje presentes mos), la cavidad craneana contiene aproximada-
en las neuronas (cap. 2), también en células de la mente 75 mL de sangre y 75 mL de líquido cefa-
glía. Tanto los oligodendrocitos como los astrocitos lorraquídeo (LCR). La función hidrostática del LCR
expresan canales de K+ dependientes del voltaje; só- es trascendente: su presencia permite la flotación
lo los astrocitos poseen canales de Na+ dependientes del cerebro, y así reduce el peso efectivo de 1.400
del voltaje. Se han identificado también distintos ti- a unos 50 gramos y sirve de amortiguación ante
pos de canales del calcio y aniónicos. Se ha propues- traumatismos craneanos.
to que estos canales son transferidos al axón, aunque
esta hipótesis no ha sido probada. La hipótesis más La mayor parte del LCR se encuentra en los ventrícu-
probable es que los canales sean operativos para los los cerebrales, donde se forma tanto por secreción des-
distintos procesos de “asistencia de la función neu- de el plexo coroideo (70%) como a partir de los capi-
ronal” regulados por la glía, ya enumerados. lares cerebrales (30%); en este último caso, el LCR
Neurociencia aplicada ©2007. Editorial Médica Panamericana
8 Parte I - Los componentes del sistema nervioso y su comunicación
Fig. 1-6. El aporte energético a la neurona está dado por la glucosa captada a través de transportadores específicos (Glut 3) y el
lactato que proviene del astrocito. El astrocito también participa en el metabolismo de transmisores (p. ej., glutamato). Hay trans-
portadores específicos de glucosa en la pared capilar y astrocito (Glut 1) y en la microglia (Glut 5, no mostrado).
Fig. 1-7. El LCR se forma y se secreta en el plexo coroideo en los ventrículos laterales, tercero y cuarto. En el adulto, el peso del
plexo coroideo es de 2-3 g. En el espacio subaracnoideo no existe plexo coroideo.
Neurociencia aplicada ©2007. Editorial Médica Panamericana
Biología de las células nerviosas 9
Fig. 1-8. Relaciones entre los componentes del espacio suba- Fig. 1-9. Meninges y espacios meníngeos. Sección coronal a
racnoideo. Espacio de Virchow-Robin. través de la región paramediana de los hemisferios cerebrales.
arriba a las cavidades ventriculares desde el espacio El término “barrera hematoencefálica” fue intro-
intersticial cerebral. Como se muestra en la figura 1- ducido por Ehrlich hacia fines del siglo XIX para de-
7, el LCR fluye desde los ventrículos laterales y nominar al fenómeno por el que una amplia gama de
a través del agujero de Monro hacia el III ventrícu- compuestos circulantes son excluidos del SNC y no
lo, y por el acueducto de Silvio, hacia el IV ven- penetran en él. Existen dos razones fundamentales
trículo. para esta exclusión: a) las características morfológi-
Desde el IV ventrículo, el LCR alcanza el espacio cas y funcionales de los capilares cerebrales y b) las
subaracnoideo por el foramen de Magendie. Dentro características fisicoquímicas de la sustancia que se
del espacio subaracnoideo, el LCR se distribuye tanto va a transferirse.
hacia abajo por el canal vertebral, como hacia arriba En los capilares cerebrales pueden distinguirse tres
por la convexidad cerebral (véase fig. 1-7). aspectos diferenciales que le dan identidad en relación
Debido a que el espacio subaracnoideo acompaña con otros capilares del organismo (fig. 1-10):
a los vasos cerebrales en trayectos prolongados den-
tro del parénquima cerebral (constituyendo los es- • El endotelio presenta uniones estrechas (tight-
pacios de Virchow-Robin), existe un pasaje fácil de junctions), que no existen en los capilares sistémi-
solutos desde el tejido cerebral hasta el espacio su- cos, y tiene muy pocas vesículas pinocitóticas. Ca-
baracnoideo y desde aquí, a los ventrículos cerebra- rece de los procesos endocitóticos (endocitosis en
les (fig. 1-8). fase fluida, endocitosis mediada por receptor) típi-
La reabsorción del LCR tiene lugar en las vellosi- cos de los capilares sistémicos.
dades subaracnoideas, que funcionan como “vál- • Las células endoteliales de los capilares cerebrales
vulas” unidireccionales del flujo (fig. 1-9). La velo- presentan numerosas mitocondrias, lo cual indica la
cidad de formación y de reabsorción del LCR es de existencia de procesos de transporte activos. En
unos 500 mL/día. El LCR y el intersticio cerebral efecto, bioquímicamente pueden demostrarse varios
están aislados de la circulación general por dos ba- mecanismos de transporte mediados por transporta-
rreras funcionales: dores (carriers) específicos, los que en muchos ca-
sos están asociados con la bomba Na+-K+-ATPasa.
• La barrera hematoencefálica, que impide el libre Los capilares cerebrales están así provistos de una
pasaje de sustancias desde los capilares cerebrales verdadera “barrera enzimática” (fig. 1-11).
al espacio extracelular del tejido nervioso. • Las células endoteliales de los capilares cerebrales
• La barrera hematocefalorraquídea, que afecta el están rodeadas (aunque no en forma total) por célu-
libre pasaje de sustancias desde los capilares coroi- las gliales, que contribuyen significativamente en
deos al LCR. dicha barrera.
Neurociencia aplicada ©2007. Editorial Médica Panamericana
10 Parte I - Los componentes del sistema nervioso y su comunicación
Fig. 1- 10. Capilares no fenestrados en el SNC. Las células en- Fig. 1-11. Procesos de transporte en el epitelio coroideo. Para
doteliales presentan “uniones estrechas” entre sí y están rodea- la secreción de LCR, tiene lugar la actividad coordinada de
das por una membrana basal y los pies de los astrocitos. transportadores de iones (círculos rojos) y canales (flechas
gruesas) en la cara basolateral (que mira al plasma) y apical
(que mira hacia el LCR). La fuerza primaria para el transporte
es la bomba Na+-K+-ATPasa; ésta mantiene la concentración de
• Puede así afirmarse que los capilares cerebrales se Na+ en las células coroideas mucho más baja que en el líquido
comportan más como órganos secretorios que como extracelular. En consecuencia, en la membrana basolateral hay
barreras de filtración. De ellos resulta la diferente una captación de Na+ hacia la célula en intercambio con H+
composición del plasma y del LCR (cuadro 1-1). (antiporte), o en el mismo sentido que el Cl- extracelular (co-
transporte). El Cl- se transporta activamente desde el plasma
Es en este nivel donde se producen los fenómenos hacia la célula a través de un antiporte y un cotransporte. En la
que conducen a la isquemia cerebral ante un daño vas- cara apical (hacia el LCR), el Na+ es bombeado activamente en
dirección de los ventrículos. A través de esta cara apical, el K+
cular. Ellos incluyen diversas manifestaciones hemo-
y el Cl–, y también el HCO3- (generado por la anhidrasa carbó-
dinámicas, electrofisiológicas y bioquímicas, con nu-
nica, c.a.), abandonan la célula a través de canales. El movi-
merosos círculos viciosos de retroalimentación positi-
miento de agua se asocia con la secreción de Cl- y K+ en el
va que amplifican el daño. La disminución del flujo LCR.
sanguíneo por debajo de cierto límite da por resultado
la disminución del aporte de O2 y una homeostasis ió-
nica alterada (salida de K+ hacia el espacio extracelu- están especializadas en la neurosecreción. Los órga-
lar y entrada de Na+ y Ca2+ en la neurona), con despo- nos circunventriculares son siete:
larización de la membrana y edema citotóxico. Se pro-
duce entonces una liberación masiva de neurotransmi- • Eminencia media del hipotálamo.
sores excitatorios (glutamato, aspartato), que es de im- • Glándula pineal.
portancia central en el establecimiento de la lesión • Órgano vasculoso de la lámina terminal.
(véase más adelante). • Área postrema.
En el SNC hay ciertas zonas (órganos circunventri- • Órgano subcomisural.
culares) donde la barrera hematoencefálica es inexis- • Órgano subfornical.
tente, debido a que los capilares carecen de las propie- • Neurohipófisis.
dades morfológicas y bioquímicas enumeradas. Los
órganos circunventriculares son verdaderas “venta- La naturaleza del compuesto que atraviesa la barrera
nas” del SNC, que cumplen funciones quimiorrecep- hematoencefálica también es de importancia para su
toras y de recepción hormonal, y que en su mayoría transferencia a través de ella. Entre las características
Neurociencia aplicada ©2007. Editorial Médica Panamericana
Biología de las células nerviosas 11
fisicoquímicas requeridas para el pasaje de compues- En la figura 1-10 se resumen las relaciones estructu-
tos en forma pasiva a través de la barrera hematoence- rales y funcionales de ambas barreras, hematoencefá-
fálica son importantes: a) un bajo peso molecular y b) lica y hematocefalorraquídea.
su afinidad por el agua, lípidos de membrana y proteí- ¿Cuál es el sitio exacto, entre los distintos compo-
nas plasmáticas y de membrana (fig. 1-12). nentes de estas barreras, en el que se ejerce la función
Las proteínas prácticamente no atraviesan en forma reguladora de la transferencia de sustancias?
pasiva la barrera hematoencefálica, mientras que en- Si bien, como ya hemos mencionado, hay zonas
tre los compuestos de bajo peso molecular, los que identificables como barreras predominantes (el endo-
son hidrosolubles la atraviesan mucho más lentamen- telio vascular para la barrera hematoencefálica; el epi-
te que los liposolubles. Hay entrada de proteínas en el telio coroideo para la barrera hematocefalorraquídea),
SNC (p. ej., citocinas) por procesos de transporte es- es más exacto considerar a las barreras como la expre-
pecífico. sión de la función conjunta de sus distintos componen-
tes, que se detallan en la figura 1-13.
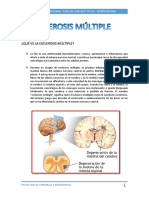
Se denomina barrera hematocefalorraquídea a Por ejemplo, en el caso de la barrera hematoencefá-
la que afecta el pasaje de sustancias desde los ca- lica, los astrocitos no forman una barrera tan continua
pilares coroideos al LCR. La barrera hematocefa- como el endotelio vascular, pero, sin embargo, sería
lorraquídea se ubica principalmente en el sello un error considerar que los astrocitos no participan en
circunferencial establecido entre las células del forma activa en el control de las sustancias que arriban
epitelio coroideo. a las neuronas desde la circulación general. Las rela-
ciones anatómicas entre estos componentes se esque-
A diferencia de los capilares cerebrales, los capilares matizan en las figuras 1-13 y 1-14.
del plexo coroideo presentan numerosas fenestracio- Las barreras hematoencefálica y hematocefalorra-
nes y, por lo tanto, su endotelio no impide la difusión quídea no están plenamente establecidas en el mo-
de sustancias desde la sangre al LCR. mento del nacimiento. Ésta es la razón por la cual cier-
Cuadro 1-1. Diferencias en concentración de diversos componentes del plasma y del LCR
Componente LCR Plasma Componente LCR Plasma
Peso específico 1,0075 1,025 Fosfato (mmol P/L) 0,48 1,3
Sólidos totales (g/100 mL) 1,0 8,7 Lactato (mEq/L) 1,7 1,7
Contenido de agua 99,0 91,3 N2 no prot (mg N/100 mL) 19 27
Sustancias reductoras 65,0 98,0 Urea 14 14
(como glucosa)
Glucosa (mg/100 mL) 61,0 92,0 Ácido úrico 0,6 1,6
No glucosa 4,0 6,0 Aminoácidos 1,6 5
Sodio (mEq/L) 141 137 Creatinina 4 6
Potasio (mEq/L) 3,3 4,9 Colesterol (mg/100/mL) 0,14 160
Calcio (mEq/L) 2,5 5,0 Proteínas (mg/100 mL) 28 7.000
Magnesio (mEq/L) 2,4 64 Albúmina 23 4.430
Base total (mEq/L) 155 163 Globulina 5 2.270
Cloro (mEq/L) 124 101 Fibrinógeno 0 300
Bicarbonato (mEq/L) 21 23
Neurociencia aplicada ©2007. Editorial Médica Panamericana
12 Parte I - Los componentes del sistema nervioso y su comunicación
Fig. 1-12. Fuerzas fisicoquímicas participantes en el pasaje de sustancias a través de la barrera hematoencefálica (BHE). AA, ami-
noácidos.
tos metabolitos circulantes, que no son nocivos duran- elementos funcionales de protección de las células
te la vida adulta para la función neuronal, lo son en la nerviosas. Su alteración, presente en diversas pa-
edad perinatal. tologías cerebrales, conlleva graves daños para la
Un ejemplo es el de la bilirrubina indirecta, que función neuronal.
cuando aumenta en el recién nacido por hemólisis ex-
cesiva (p. ej., incompatibilidad Rh) produce un cuadro En el cuadro 1-2 se enumeran algunas propiedades
de daño irreversible de los ganglios basales llamado de la barrera hematocefalorraquídea.
“kernicterus”. En cambio, en los adultos, ictericias
aún más pronunciadas por bilirrubina directa no cau- El cerebro está protegido por una
san daño cerebral debido a la existencia de las barre-
ras ya mencionadas y a la menor toxicidad del com-
estructura indeformable de hueso
puesto en forma conjugada. craneano
En conclusión, las barreras hematoencefálica y El flujo sanguíneo cerebral en un adulto normal es de
hematocefalorraquídea deben considerarse como 750 mL/min (50 mL/100 g/min); a la sustancia gris le
Neurociencia aplicada ©2007. Editorial Médica Panamericana
Biología de las células nerviosas 13
Cuadro 1-2. Propiedades de la barrera hematocefalorraquídea
Generador de agua Transporte activo desde SNC
Sitio principal de pasaje de agua de sangre a SNC. LCR co- Aniones orgánicos (p. ej., ácido homovanílico) y iones inor-
mo amortiguador para traumatismos encefálicos gánicos (p. ej., yoduro) son eliminados activamente mante-
niendo bajos los niveles de LCR
Proveedor de micronutrientes Metabolismo de fármacos
Extracción y transporte de nucleósidos, microelementos, vita- El plexo coroideo metaboliza, como el hígado, xenobióticos
minas hidrosolubles, etc. Desde la sangre al LCR para entrar que poseen enzimas como P-450 o epóxido hidrolasa
en neuronas o en la glía
Fábrica de factores tróficos Vigilancia inmunológica
Producción y secreción al LCR de IGF-I, TF-β, etc. Debido a la falta de linfáticos del tejido nervioso, las células
presentadoras de antígenos interaccionan en el plexo coroi-
deo con los linfocitos
Transporte de fármacos Fuente o blanco de neuropéptidos
Se puede evitar la BHE usando los sistemas de transportado- Receptores y/o síntesis de vasopresina, insulina, angiotensina
res del plexo coroideo (p. ej., AZT) II, leptina. Estos péptidos actúan localmente o se distribuyen
en el tejido cerebral
Homeostasis iónica del LCR Entrada de agentes patógenos
Sensado de cambio en iones K+, Ca2+ y Mg2+ en el LCR y Debido a la permeabilidad del plexo coroideo, los complejos
ajuste de la velocidad de transporte a ellos. Regulación del inmunes o gérmenes pueden atravesarlo (p. ej., HIV)
transporte de Cl- y HCO3- para mantener el pH del LCR
Fig. 1-13. Relaciones funcionales entre los distintos elementos
que componen las barreras hematoencefálica y hematocefalo-
rraquídea. Las flechas indican la dirección del flujo del LCR.
(Modificado de Kandel y col, 2000.)
Neurociencia aplicada ©2007. Editorial Médica Panamericana
14 Parte I - Los componentes del sistema nervioso y su comunicación
Fig. 1-14. Células participantes en el intercambio entre compartimientos cerebrales. (Modificado de Kandel y col, 2000.)
corresponden 75 mL/100 g/min y a la sustancia blanca, La mayor parte de la energía cerebral es
25 mL/100 g/min. Como hemos dicho, la presencia de consumida para el mantenimiento del
LCR reduce el peso efectivo del cerebro. Esto, junto con gradiente iónico
la rigidez de la estructura ósea craneana, aumenta la pro-
tección del SNC ante el trauma, pero lo hace susceptible, Entre el 50% y el 80% del metabolismo energé-
ante un desequilibrio del contenido del cráneo, a un au- tico cerebral se invierte en el trabajo de la bomba
mento de la presión intracraneana. Na+-K+-ATPasa, mientras que la biosíntesis de neu-
rotransmisores sólo insume un 1% del total.
El componente principal que ocupa la cavidad
craneana es el agua, distribuida en cuatro com- El resto de la energía se utiliza en tareas de biosínte-
partimientos: sangre, LCR y los espacios extracelu- sis neuronal (renovación de membranas celulares y la
lar e intracelular (neuronal y glial). En forma síntesis de proteínas estructurales y enzimas). Debe
esquemática, puede decirse que el 80% del conte- notarse que existe un estrecho acoplamiento funcional
nido intracraneano está constituido por la masa entre el metabolismo cerebral, la actividad neuronal y
encefálica, el 10% por la sangre de los vasos san- el flujo sanguíneo cerebral.
guíneos y un 10% por el LCR. Ante incrementos de la actividad neuronal y de la
demanda metabólica cerebral, se produce, por acción
Para su integridad estructural y funcional, el cerebro de quimiorreceptores vasculares, un incremento del
depende del aporte constante de glucosa y oxígeno y de flujo sanguíneo cerebral. Este acoplamiento tiene
la eliminación de sus desechos metabólicos. Esto impli- una latencia de unos 2 segundos y es estrictamente re-
ca una íntima relación entre el flujo sanguíneo cerebral, gional.
la disponibilidad de los sustratos necesarios y los reque- Depende principalmente de la acción de señales que
rimientos metabólicos cerebrales (fig. 1-15). se acumulan en el líquido extracelular durante la acti-
Neurociencia aplicada ©2007. Editorial Médica Panamericana
Biología de las células nerviosas 15
Fig. 1-15. Factores que afectan el flujo sanguíneo cerebral. (Modificado de Ganong y col, 2003.)
vación neuronal, como lactato, H+, adenosina, K+, neo cerebral. Así, los incrementos de la actividad fun-
prostaglandinas, óxido nítrico (NO) y, en forma secun- cional cerebral están asociados con aumentos del flu-
daria, de la acción de neurotransmisores sobre recep- jo sanguíneo cerebral, lo cual da la base para la mayo-
tores en la microcirculación cerebral, como la nora- ría de los métodos de neuroimagen funcional en uso
drenalina, la acetilcolina, el péptido vasoactivo intesti- en la actualidad.
nal (VIP) o la sustancia P. El efecto del O2 es de menor cuantía. Sólo cuando la
En condiciones basales, la utilización celular de glu- presión parcial de O2 cae por debajo de 50 mm Hg se
cosa, el consumo de oxígeno y el flujo sanguíneo ce- produce vasodilatación.
rebral están en estrecha relación. Cuando aumenta la Otros mecanismos que mantienen la perfusión cere-
actividad sináptica (liberación de neurotransmisor), bral normal son la vasodilatación refleja (manteni-
aumentan los requerimientos metabólicos (a través de miento de un flujo normal mediante la reducción de la
la glucólisis) para el metabolismo de neurotransmiso- resistencia vascular), la circulación por arterias colate-
res (en especial en los astrocitos, que recaptan al neu- rales y el incremento en la cantidad de extracción ce-
rotransmisor glutamato para procesarlo a glutamina) rebral de glucosa y O2.
(véase fig. 1-6). Debe notarse que el control neurogénico de la circu-
No es de extrañar entonces que el CO2 sea el agente lación cerebral no tiene un papel tan importante en la
fisiológico y farmacológico más potente para modifi- regulación del flujo sanguíneo cerebral como los fac-
car el flujo sanguíneo cerebral. Los vasos cerebrales tores metabólicos antes mencionados. El sistema ner-
reaccionan casi instantáneamente ante cambios en la vioso autónomo simpático cervical (proveniente del
presión local de CO2. Su aumento genera vasodilata- ganglio cervical superior) provee vasoconstricción no-
ción y su descenso tiene el efecto contrario. radrenérgica a las grandes arterias cerebrales, mientras
Un cambio de 1 mm Hg en la presión parcial arterial que el parasimpático cerebral es vasodilatador por ac-
de CO2 produce un aumento del 2% en el flujo sanguí- ción de la acetilcolina en ese mismo nivel. El tono va-
Neurociencia aplicada ©2007. Editorial Médica Panamericana
16 Parte I - Los componentes del sistema nervioso y su comunicación
soconstrictor de la microcirculación depende de la cerebral constante ante cambios de la presión arterial
actividad de neuronas noradrenérgicas del locus coe- sistémica, y así se mantiene la provisión constante de
ruleus y serotoninérgicas del rafe. Hay también inter- O2 y glucosa para las células cerebrales. El aumento
neuronas corticales peptidérgicas que usan neuropép- de CO2 , y la caída del pH y de la PO2, inducen la for-
tido Y para promover la vasoconstricción o VIP para la mación de NO en la pared vascular, lo cual causa rela-
vasodilatación. jación vascular. La caída de CO2, y el aumento del pH
y de la PO2 producen vasoconstricción y aumento de
La autorregulación vascular cerebral la resistencia vascular.
previene que ocurran modificaciones
importantes en el flujo sanguíneo En la isquemia cerebral se compromete
cerebral ante cambios sistémicos el flujo sanguíneo y disminuyen el
aporte de O2 y glucosa y la eliminación
En condiciones normales, el flujo sanguíneo cerebral de productos del catabolismo cerebral
se mantiene constante a través de un amplio rango de
variación de la presión de perfusión cerebral (dada por El cerebro tiene depósitos mínimos de energía, por
la diferencia entre la presión arterial media y la pre- lo que la lesión por isquemia es mayor que en otros
sión intracraneana y cuyo valor normal varía entre 5 y tejidos (figs. 1-16 a 1-18, recuadro 1-1.). La isquemia
20 cm de agua). Por este mecanismo de autorregula- global se produce por caída de la presión arterial sis-
ción vascular cerebral se previene que cambios sisté- témica o por aumento de la presión intracraneana. Una
micos generen modificaciones importantes del flujo isquemia global de 5 a 10 minutos produce daño per-
sanguíneo cerebral. manente e irreversible de las células nerviosas.
La autorregulación resulta de un mecanismo miogé- ¿Cómo cambia la microcirculación en la isquemia?
nico controlado por la presión intraluminal (su aumen- La disminución de nutrientes y el aumento de produc-
to produce vasoconstricción y su disminución, vasodi- tos de desecho son señales de aumento del flujo san-
latación) y que opera en forma independiente y simul- guíneo local para mantener la presión de perfusión.
tánea con los otros factores neurogénicos, químicos y Los vasos se dilatan para reducir la resistencia, la pre-
metabólicos. sión arterial aumenta para mantener la perfusión, y
La autorregulación cerebral mantiene el flujo cons- existen factores locales de resorción del coágulo (véa-
tante ante modificaciones en la presión de perfusión se fig. 1-17). La isquemia depleciona las reservas
entre 60 y 150 mm Hg. Esto protege al SNC, por energéticas. No hay energía para mantener los gra-
ejemplo, de los cambios posturales, de las eventuales dientes de concentración de Na+ y K+, las neuronas se
oclusiones arteriales o del aumento de la presión intra- despolarizan y se liberan neurotransmisores. Se dañan
craneana. las mitocondrias y se afecta la cadena respiratoria, con
Es de notar que existe un acoplamiento efectivo en- producción de radicales libres. La glucosa se convier-
tre la presión intracraneana y la presión arterial sisté- te en lactato con reducción de la producción de ATP.
mica. Ante el aumento de la presión intracraneana, au- En la isquemia central se produce un área periférica
menta la presión venosa intracerebral y disminuye el de “penumbra” (véase fig. 1-16). El destino de esta
flujo sanguíneo cerebral. Esto genera en forma refleja zona indefinida (muerte o recuperación) dependerá de
un aumento de la presión arterial sistémica (reflejo de la rapidez y eficacia de las medidas médicas adoptadas
Cushing). en la fase aguda de la isquemia.
En síntesis, puede decirse que la irrigación del cere- En la isquemia se abren canales iónicos en la mem-
bro depende de la presión de perfusión, o sea, de la di- brana celular de las neuronas, el Na+ y el H2O entran
ferencia entre la presión arterial sistémica media y la en la célula y causan edema celular. Hay liberación de
presión intracraneana. La presión de perfusión puede glutamato y reducción de su captación neuronal y glial
caer por: por menor disponibilidad de ATP. El glutamato se une
a receptores NMDA y no NMDA (véase cap. 3) con
• Disminución del volumen sistólico. entrada de Ca2+ en las células. El aumento de Ca2+
• Incremento de la presión intracraneana. produce lesión neuronal, liberación de fosfolipasas y
• Vasoconstricción local. alteración de fosfolípidos de membrana, con forma-
ción de eicosanoides, prostaglandinas, tromboxanos y
La resistencia local se controla por factores metabó- leucotrienos. Esto lleva a mayor vasoconstricción,
licos locales (autorregulación), que mantienen el flujo edema y coagulación intravascular. El aumento de
Neurociencia aplicada ©2007. Editorial Médica Panamericana
Biología de las células nerviosas 17
Fig. 1-16. Evolución de la zona de penumbra luego de la isquemia cerebral.
Fig. 1-17. Fenómenos celulares en la isquemia cerebral.
Neurociencia aplicada ©2007. Editorial Médica Panamericana
18 Parte I - Los componentes del sistema nervioso y su comunicación
RECUADRO 1-1
Accidentes cerebrovasculares (I)
Los accidentes cerebrovasculares son una de las tres teral alcanza a compensar la reducción. El estrecha-
causas más frecuentes de coma cerebral y muerte. Con- miento arterial es producido en general por depósitos
sisten en la disfunción neurológica producida por la re- de lípidos en la pared (ateromas).
ducción del flujo sanguíneo cerebral. El cuadro neuro- Una caída de la presión sistémica severa puede con-
lógico puede ser transitorio o definitivo. La isquemia ducir a una disminución del flujo cerebral, aun en
cerebral es una alteración potencialmente reversible presencia de vasos normales. Esta situación origina in-
de la función cerebral, resultante de la provisión inade- fartos en las zonas de borde, es decir, en las áreas loca-
cuada de oxígeno o glucosa. Si la isquemia es grave co- lizadas entre la distribución de dos arterias mayores.
mo para producir muerte celular, se llega al infarto ce- Como estas zonas están al final de ambos árboles arte-
rebral, situación en que las posibilidades de reversión riales, están sujetas a una perfusión sanguínea baja,
disminuyen considerablemente. La muerte neuronal que en condiciones normales es sólo marginalmente
sobreviene a los 5-10 minutos de isquemia. suficiente. Son, por lo tanto, las primeras zonas en com-
La falla en la disponibilidad de energía por las células prometerse ante caídas de la presión arterial sistémica.
cerebrales es la base de los síntomas neurológicos del Si esta caída es prolongada y de importancia, sobrevie-
accidente cerebrovascular. La muerte neuronal se pro- ne una isquemia cerebral global.
duce cuando las neuronas son incapaces de sintetizar La hemorragia cerebral es una de las formas más
ATP. Al no contar con nutrientes, la supervivencia celu- graves de accidente cerebrovascular y resulta de la
lar se compromete. Hemos visto en este capítulo que ruptura espontánea de la pared de un vaso sanguíneo
como resultado de la acidosis intracelular por la glucó- debilitado por una hipertensión arterial de larga evolu-
lisis anaeróbica se deprime la respiración mitocondrial, ción, o por la presencia de un ensanchamiento congé-
se producen radicales libres y tiene lugar una intensa nito de la pared o un aneurisma. En el primer caso, la
peroxidación de lípidos. También se altera la homeos- hemorragia ocurre hacia el parénquima cerebral (he-
tasis iónica neuronal con entrada de Na+, Cl-, H2O y, so- morragia intracerebral). En el segundo caso, se acom-
bre todo, de Ca2*. La entrada de agua conduce al ede- paña además de hemorragia hacia el LCR, dado que los
ma celular, con compresión de los vasos sanguíneos y aneurismas se ubican en general en la superficie de los
mayor reducción de la circulación. Las estructuras celu- hemisferios. Ambos tipos de hemorragias (intracere-
lares se degeneran porque no existe la energía necesa- bral, subaracnoidea) son de pronóstico serio, debido al
ria para la síntesis de macromoléculas. efecto de masa y compresión de estructuras cerebrales
Otro factor agravante es la pérdida de los mecanis- vecinas y al severo espasmo de los vasos cerebrales de-
mos de autorregulación del flujo cerebral, discutidos en bido a la presencia de sangre en el LCR.
este capítulo. Como vimos, este proceso mantiene rela- Los accidentes cerebrovasculares están en general pre-
tivamente constante el flujo cerebral a pesar de las va- cedidos por ataques isquémicos transitorios, que ceden
riaciones de la presión arterial media. El sistema es efi- en forma espontánea (en un lapso de 15 minutos a 24
caz hasta un nivel inferior de presión arterial media de horas). Una causa común de estos ataques transitorios es
60 mm Hg, con límite superior en los 150 mm Hg. En el breve episodio de isquemia producido por el pasaje de
el área de infarto cerebral, la autorregulación desapare- un émbolo, que produce obstrucción hasta que el émbo-
ce y el flujo sanguíneo sigue entonces en forma pasiva lo se destruye y fluye por el árbol circulatorio. Estos ém-
a los cambios en la presión arterial sistémica. bolos pueden originarse en el corazón o en una lesión
El flujo sanguíneo cerebral disminuye ante cualquier arteriosclerótica de un vaso grande, como la carótida. Un
proceso que estreche u ocluya un vaso cerebral nu- sitio común es la bifurcación de ésta en su rama interna
triente. Se llama estenosis a la oclusión parcial. En el y externa. Los émbolos producidos pueden causar disfun-
caso de la carótida, se requiere una reducción del 50% ción sensorial, motora o del lenguaje, o ceguera unilate-
al 75% del diámetro antes de que haya modificación ral transitoria. Los ataques isquémicos transitorios deben
severa del flujo. Aun en estas circunstancias, el flujo ce- evaluarse y diagnosticarse cuidadosamente a fin de pre-
rebral puede permanecer normal si la circulación cola- venir episodios de mayor gravedad.
Ca2+ activa la producción de radicales libres que se con aumento de la permeabilidad de la barrera hema-
difunden a otras neuronas alterándolas con destruc- toencefálica. Así, proteínas del plasma entran en el es-
ción celular (véase fig. 1-18). pacio intersticial cerebral y se produce edema vasogé-
Otros factores agravantes son el edema de astrocitos nico con aumento de la presión intracraneana y mayor
perineuronales y perivasculares y el daño endotelial compromiso del flujo sanguíneo. El cuadro clínico
Neurociencia aplicada ©2007. Editorial Médica Panamericana
Biología de las células nerviosas 19
Fig. 1-18. Progresión de fenómenos en la isquemia cerebral.
que se produce es altamente dependiente del territorio negativo (unos -60 a -70 mV). Este fenómeno no es
vascular involucrado (recuadro 1-2). privativo de las neuronas. Los valores del potencial de
reposo en distintas células del organismo varían entre
Las neuronas presentan un potencial -40 y -75 mV, con excepción del músculo esqueléti-
de reposo y cuatro tipos de señales co, donde alcanza unos -90 mV. Cuando el potencial
de reposo de la membrana se hace más negativo que
eléctricas en la situación de reposo, es decir, cuando aumenta,
se habla de hiperpolarización. Por el contrario, una
Las señales neurales dependen de las propiedades reducción en el potencial de membrana, por ejemplo,
eléctricas de la membrana celular, y en las neuronas se de –70 a –40 mV, se llama despolarización. La hiper-
observan distintos tipos de potenciales. polarización hace a la neurona menos excitable,
En forma general, y con dependencia de la región mientras que la despolarización la transforma en más
neuronal examinada, las neuronas presentan un poten- excitable.
cial de reposo y las siguientes señales eléctricas:
La señal de entrada comprende dos variantes,
• Señal de entrada. según se trate de la superficie receptora de las
• Señal de integración. neuronas sensoriales o de las superficies dendrí-
• Señal de conducción. tica o somática de las neuronas centrales. En las
• Señal de salida o de secreción (fig. 1-19). neuronas sensoriales, el cambio de potencial se
denomina potencial receptor o generador; en
El potencial de reposo resulta, como en toda célula las dendritas o el soma neuronal se llama poten-
del organismo, de la separación de cargas eléctricas a cial sináptico.
través de una membrana celular que es semipermea-
ble. Si el valor del potencial extracelular se fija en for- Ambos potenciales son de naturaleza local, gradua-
ma arbitraria en 0 mV, el interior de las neuronas será dos y de propagación pasiva o electrotónica; disminu-
Neurociencia aplicada ©2007. Editorial Médica Panamericana
20 Parte I - Los componentes del sistema nervioso y su comunicación
RECUADRO 1-2
Accidentes cerebrovasculares (II)
El déficit neurológico producido por los accidentes de la homónima del hemisferio opuesto, el cuadro con
cerebrovasculares depende del vaso sanguíneo involu- frecuencia se limita al compromiso del territorio de la
crado. El cerebro está perfundido por las arterias caró- cerebral media (véase fig. 1-21)
tidas y las basilares (figs. 1-20 y 1-21). Uno de los cua- La oclusión de una arteria vertebral puede pasar
dros más comunes involucra al territorio de la arteria inadvertida si la vertebral opuesta está normal y aporta
cerebral media. Esta arteria tiene dos ramas: una pro- circulación colateral a través de la arteria basilar. En
funda (la lenticuloestriada) y otra superficial (pial). La otros casos, la oclusión de la arteria vertebral puede de-
rama profunda irriga la cápsula interna, parte del globo rivar en infarto del territorio de unas de sus ramas, la ar-
pálido y del caudado, y la corona radiata. La rama pial teria cerebelosa posteroinferior. Esto desencadena un
irriga la superficie lateral de los lóbulos frontal, tempo- cuadro de compromiso de la porción lateral del bulbo,
ral y occipital. El cuadro clínico que resulta de la este- conocido como “síndrome de Wallenberg”. Las estruc-
nosis u oclusión de la arteria cerebral media depende turas afectadas son la rama espinal del trigémino, el
de cuál de las ramas es la más afectada. Entre los sín- tracto espinotalámico, el núcleo ambiguo del vago,
tomas más comunes se encuentran la hemiparálisis y la el pedúnculo cerebeloso inferior y las fibras simpáticas
pérdida de sensibilidad contralateral, ambas más pro- descendentes (cap. 12). Como consecuencia, el síndro-
nunciadas en el miembro superior. Esto se debe a que me de Wallenberg comprende, desde el punto de vista
la representación del miembro inferior en la corteza sensorial, pérdida de la sensibilidad dolorosa y térmica
sensorial y motora primaria está en la superficie medial (pero no táctil) de la porción homolateral de la cara (vía
de los lóbulos frontal y parietal (“hombrecillo invertido no cruzada del tracto espinal del V par) y pérdida de la
con los miembros inferiores colgando el espacio inter- sensibilidad al dolor y temperatura de la mitad opues-
hemisférico”; cap. 4). Estas áreas están fuera del territo- ta del cuerpo (por lesión de la vía espinotalámica, que
rio de la cerebral media. es cruzada) (cap. 4). Hay incoordinación homolateral
La afasia es común en las lesiones vasculares del he- de los miembros (por lesión del pedúnculo cerebeloso
misferio dominante (cap. 16). Cuando la lesión ocurre inferior) y disfonía (por parálisis homolateral de las
en el hemisferio no dominante, en especial en el lóbu- cuerdas vocales (lesión del núcleo ambiguo del X par).
lo parietal, se produce una alteración grave de la repre- En el ojo homolateral se observan ptosis (caída del pár-
sentación espacial (abandono del hemicuerpo contra- pado) y miosis (constricción de la pupila) por lesión del
lateral o neglect syndrome), en el cual el paciente no simpático. Se alteran también de manera significativa
atiende a objetos o estímulos localizados contralateral- los mecanismos del sueño y del soñar (cap. 15). El sín-
mente a la lesión (cap. 16). En forma independiente de drome de Wallenberg es un buen ejemplo de correla-
este cuadro, puede presentarse hemianopsia contrala- ción anatómica, fisiológica y clínica. También ilustra un
teral cuando están involucradas las radiaciones ópticas, hecho de interés: cuando existen cuadros sensoriales o
es decir, las vías talamocorticales que conectan el cuer- motores cruzados (un lado de la cara y el lado opuesto
po geniculado lateral con la corteza visual (cap. 5). corporal), ello implica lesiones del tronco encefálico.
La arteria cerebral anterior irriga al lóbulo frontal La obstrucción de la arteria basilar lleva al infarto
anterior y a partes de la corteza frontal y parietal en la de la porción superior del tronco encefálico y de ambos
región interna de los hemisferios. Por las razones ya ci- lóbulos occipitales. Este cuadro con frecuencia es fatal.
tadas, la alteración del flujo en esta arteria se acompa- El par de arterias cerebrales posteriores se origina de la
ña de parálisis y de pérdida de la sensibilidad en el bifurcación de la porción terminal de la arterial basilar.
miembro inferior contralateral. Este cuadro no se acom- Cada arteria cerebral posterior tiene una rama hemisfé-
paña de hemianopsia ni de afasia. Los ojos pueden es- rica que irriga al lóbulo occipital y ramas perforantes
tar desviados hacia el sitio de la lesión debido al com- que irrigan al tronco encefálico, junto con otras ramas
promiso del área frontal de la mirada, responsable de de la basilar (véase fig. 1-20). La oclusión de las ramas
dirigir los movimientos oculares rápidos de persecución hemisféricas de una arteria cerebral posterior produce
de objetos en el plano horizontal (véase cap. 5). Cuan- hemianopsia (pérdida de la mitad del campo visual)
do esta zona está dañada, predomina la del hemisferio contralateral homónima en ambos ojos (cap. 5). Por
opuesto, razón por la que el enfermo tiene los ojos des- ejemplo, una oclusión de la arteria cerebral posterior
viados hacia la lesión (véase fig. 10-24). derecha produce un infarto occipital derecho con pér-
La oclusión de la arteria carótida interna da por re- dida de la mitad izquierda del campo visual de ambos
sultado el infarto de los dos tercios anteriores del he- ojos.
misferio correspondiente, en el área de distribución de Cuando se ocluyen ambas ramas hemisféricas de las
las dos arterias mencionadas, cerebral media y anterior. cerebrales posteriores, la pérdida total de la visión que
Como la arteria cerebral anterior recibe flujo colateral se produce se denomina “ceguera cortical” (cap. 5). Los
Neurociencia aplicada ©2007. Editorial Médica Panamericana
Biología de las células nerviosas 21
RECUADRO 1-2 (Cont.)
Accidentes cerebrovasculares (II)
Fig. 1-20. Irrigación del cerebro, vista basal. La vía sanguínea principal es a través de la arteria carótida interna y el sistema
vertebrobasilar, los que se comunican entre sí a través del polígono de Willis.
Fig. 1-21. Superficies lateral y medial del cerebro, que muestran la distribución de las principales arterias cerebrales. Las ar-
terias cerebral anterior y media son ramas de la carótida interna; la arteria cerebral posterior es rama de la arteria basilar.
Neurociencia aplicada ©2007. Editorial Médica Panamericana
22 Parte I - Los componentes del sistema nervioso y su comunicación
RECUADRO 1-2 (Cont.)
Accidentes cerebrovasculares (II)
pacientes con este cuadro muchas veces niegan la sia, se afecta la visión de color del hemicampo visual
existencia de la ceguera. opuesto a la lesión en ambos ojos.
Cada arteria cerebral posterior también perfunde al Además de las lesiones citadas, que corresponden a
esplenio, denominación que recibe la porción posterior las grandes ramas de las arterias cerebrales, se produ-
del cuerpo calloso. Cuando esta estructura se infarta, cen también infartos lacunares, o pequeñas lesiones
en conjunto con la corteza visual primaria del hemisfe- de menos de 15 mm de diámetro, debidas a la oclu-
rio dominante, se origina un cuadro de alexia (imposi- sión de arterias penetrantes pequeñas que se han alte-
bilidad de comprender la palabra escrita) sin agrafia rado por la hipertensión crónica. Aunque de poca ex-
(imposibilidad de escribir) (véase cap. 16). tensión, estas lesiones suelen ser devastadoras. Por
El cuadro de hemiacromatopsia (pérdida de la visión ejemplo, un infarto lacunar que implica a la cápsula in-
cromática de un hemicampo visual) se origina cuando se terna o al tracto piramidal en la protuberancia puede
lesionan las áreas secundarias visuales en el infarto de la producir una hemiparesia grave, o un infarto lacunar
porción inferior y medial del lóbulo occipital. Esto se de- del núcleo ventral posterior del tálamo puede producir
be a que en estas áreas se encuentran zonas que discri- una pérdida sensorial severa contralateral con síndro-
minan el color (cap. 5). Como en el caso de la hemianop- me talámico (cap. 4).
Fig. 1-19. Las distintas señales de recepción, integración, conducción y secreción en neuronas sensoriales, motoras e interneuro-
nas. A la derecha, los distintos potenciales encontrados en cada segmento. (Modificado de Kandel y col. 2000)
Neurociencia aplicada ©2007. Editorial Médica Panamericana
Biología de las células nerviosas 23
yen en forma progresiva en intensidad, y no se detec- interneurona). La amplitud del potencial de acción
tan más allá de 1 o 2 mm del sitio de origen. Su ampli- es de unos 100 mV y dura 0,5-2 mseg.
tud es de 0,1 a 5 mV, excepto en casos particulares co-
mo la placa motora (véase cap. 3) o en las sinapsis de la La señal de salida se observa en las terminales si-
fibras trepadoras con células de Purkinje del cerebelo nápticas del axón, donde la despolarización produce la
(véase cap. 11). Los potenciales receptores o generado- liberación de neurotransmisor (sinapsis de tipo quími-
res se detectan en los receptores sensoriales y son, en sus co) o perturba, debido a la aposición de membranas, el
distintas variantes, una representación analógica del es- potencial de reposo de la neurona postsináptica (si-
tímulo. Pueden ser hiperpolarizantes (inhibitorios) o napsis de tipo eléctrico).
despolarizantes (excitatorios).
Los potenciales sinápticos son el medio por el cual En el caso de las sinapsis químicas, la libera-
una neurona puede modificar el potencial de membra- ción de transmisor depende de la entrada de
na de las células con las cuales se conecta. Para ello, Ca2+ e implica la generación de un potencial
la neurona presináptica libera un transmisor químico local, llamado potencial secretor, desencade-
o, con menor frecuencia, la transmisión se realiza por nado por el potencial de acción.
un mecanismo eléctrico.
En la transmisión química, el neurotransmisor inte- La entrada de Ca2+ es proporcional a la intensidad
ractúa con receptores ubicados en la superficie de la del potencial secretor y es esencial para la liberación
membrana postsináptica, lo cual da lugar a la genera- exocitótica del transmisor.
ción del potencial sináptico, que puede ser de tipo in- La distribución de canales dependientes del voltaje
hibitorio: potencial inhibitorio postsináptico (PIPS) señalada (de Na+ y K+ en el axón; de Ca2+ en la termi-
(que es hiperpolarizante) o excitatorio: potencial ex- nal neural) no debe tomarse como absoluta. En las
citatorio postsináptico (PEPS) (de naturaleza despo- dendritas coexisten los tres tipos de canales depen-
larizante). La duración de los potenciales sinápticos es dientes del voltaje en regiones intersinápticas de la
variada (desde milisegundos a, en ciertos casos, se- membrana celular; también están presentes los canales
gundos o minutos). regulados por transmisor, característicos de la región
La señal de integración se observa en la “zona gati- sináptica. Esta coexistencia de canales de distintos ti-
llo” de la membrana neuronal, donde los distintos poten- pos define el perfil de descarga típico de cada neurona
ciales locales, propagados electrotónicamente, se suman (véase cap. 3).
y dan origen al potencial de acción. En general, aunque
no siempre, la “zona gatillo” se ubica en el cono axonal. Cada neurona comprende un conjunto
Esta zona se caracteriza por poseer una concentración
de macromoléculas específicas y no
elevada de canales de Na+ y K+ dependientes del volta-
je, particularidad que la transforma en la porción de me- específicas
nor umbral de toda la membrana celular. Si la suma de
los potenciales sinápticos alcanza el umbral, se genera Hemos mencionado que las formas neuronales son
un potencial de acción; de allí que se llame “integrado- en extremo variadas (unas 10.000). Esta diversidad ci-
ra” a la señal producida. Veremos en el siguiente capítu- tológica es el resultado del proceso embriológico co-
lo que dicha suma puede ser de tipo espacial o temporal. nocido con el nombre de diferenciación. Cada célula
La señal de conducción es el potencial de acción. diferenciada sintetiza sólo ciertas macromoléculas
Mientras que los potenciales sináptico o receptor se (enzimas, proteínas estructurales, componentes de
propagan en forma pasiva y disminuyen en amplitud membrana, productos de secreción), es decir, utiliza
con la distancia, el potencial de acción (o “potencial sólo una porción del material genético que contiene.
espiga”) tiene las siguientes propiedades: Muchos componentes de las neuronas son comunes
a otras células y, por lo tanto, no son específicos. Otros
• Se propaga activamente a lo largo del axón (o en componentes se encuentran sólo en las neuronas, o
ciertos casos, como las neuronas piramidales de la únicamente en ciertos grupos neuronales, y son enton-
corteza cerebral, también por las dendritas). ces específicos. Es decir, cada neurona comprende
• No disminuye su intensidad en función de la distan- un conjunto de macromoléculas específicas y no espe-
cia. cíficas.
• Es de naturaleza “todo o nada”. Como ejemplo de lo antedicho, mencionamos aquí
• Es semejante en todas las neuronas, sea cual fuere la algunas diferencias y semejanzas entre los dos compo-
función que tenga la neurona (sensorial, motora o de nentes neuronales del reflejo miotático, cuya función
Neurociencia aplicada ©2007. Editorial Médica Panamericana
24 Parte I - Los componentes del sistema nervioso y su comunicación
Fig. 1-22. Estructura histológica de una motoneurona del asta anterior de la médula espinal. A. Un único axón mielinizado se ex-
tiende desde el asta anterior medular a las fibras musculares. B. Sección transversal a través de las porciones internodales que com-
prenden las capas de mielina formadas por la célula de Schwann. C. Sección longitudinal del nodo de Ranvier, con en axón cen-
tral desprovisto de la capa de mielina.
se analiza en detalle en el capítulo 9 (véase fig. 9-14). aferentes primarias, acetilcolina para las motoneu-
El reflejo miotático está mediado por una neurona ronas alfa).
sensorial primaria aferente (Ia), con su soma ubica-
do en los ganglios de las raíces dorsales, y dos prolon- Como semejanzas entre ambas neuronas pueden
gaciones, una periférica que termina en el huso mus- mencionarse, entre otras propiedades:
cular del músculo esquelético, y una central hacia la
médula espinal. El segundo componente neuronal de • Canales similares de Na+, K+ y Ca2+ dependientes
este reflejo es la motoneurona alfa ubicada en el as- del voltaje en la membrana neuronal.
ta anterior de la médula espinal, y sobre la cual hace • Tienen un idéntico mecanismo de intercambio Na+
sinapsis la prolongación central de la aferente prima- -K+ (la bomba Na+-K+-ATPasa).
ria Ia. La neurona sensorial primaria y la motoneuro- • Ambos tipos de neuronas presentan axones envuel-
na alfa difieren entre sí en: tos por una vaina de mielina (fig. 1-22).
• Su forma (seudounipolar en las aferentes primarias, Es decir, las similitudes y las diferencias dependen de
multipolar en el caso de las motoneuronas alfa). la síntesis y la distribución de las proteínas neuronales.
• En el tipo de conexiones que recibe (la información La fracción de material genético expresada por las cé-
de entrada llega a la motoneurona a nivel de las den- lulas nerviosas es la mayor del organismo. Se calcula que
dritas en un 95% y sólo el 5% en el cuerpo neuro- unas 200.000 secuencias distintas de RNA mensajero son
nal; en el caso de las neuronas sensoriales, ello ocu- expresadas en el cerebro, lo cual constituye unas 10-20
rre en uno de los extremos seudounipolares). veces más que lo observado en el hígado o el riñón. La
• En el tipo de receptor presente en sus membranas velocidad de expresión de estos genes es variada. Los es-
celulares (sensible a la deformación celular produci- tudios sobre genes de expresión temprana (p. ej., onco-
da por el estiramiento del músculo en las aferentes gén c-fos) han incorporado un elemento dinámico en la
primarias; específico para neurotransmisores como descripción de las conexiones cerebrales, ya que se con-
el glutamato, el GABA y la glicina en las motoneu- sideran marcadores de la actividad neuronal. En este sen-
ronas alfa). tido, los resultados obtenidos coinciden con los de la au-
• En el transmisor que emplean (glutamato para las torradiografía con glucosa radiactiva (véase cap. 10). Un
Neurociencia aplicada ©2007. Editorial Médica Panamericana
Biología de las células nerviosas 25
adelanto de interés es el análisis mediante el desarrollo de lo endoplasmático, que luego se transferirán, previo
formas atenuadas de virus (herpes simple, adenovirus) pasaje por el aparato de Golgi, a la membrana celu-
que infectan a las neuronas y permiten la transferencia de lar (proteínas de membrana) o a distintas organelas,
genes a las neuronas maduras adultas. Así se puede indu- como la membrana nuclear, el aparato de Golgi, las
cir la síntesis de proteínas que desempeñan un papel crí- vesículas secretorias, los endosomas, o el mismo re-
tico en la fisiología neuronal. Esta manipulación genéti- tículo endoplasmático. Existen varias configuracio-
ca es específica bioquímica y anatómicamente, y puede nes de inserción de proteínas a membranas, según la
realizarse en regiones individualizadas del encéfalo adul- atraviesen por un único sitio de inserción o varios
to. Abre también la posibilidad de la terapia génica. Con (ejemplo de este último caso son las proteínas cons-
excepción de algunas pocas proteínas codificadas por el titutivas de los canales iónicos).
genoma mitocondrial, todas las especies de RNA mensa- • Se trasloca a la luz de las cisternas del retículo (pro-
jero en las neuronas tienen origen nuclear. teínas secretorias). En el caso de las proteínas secre-
Las neuronas, como otros tipos de células, sintetizan torias, durante este período se produce un procesa-
tres clases de proteínas: do activo del péptido original, que incluye ruptura
de la proteína en fragmentos de menor peso molecu-
• Proteínas que se sintetizan en el citoplasma y per- lar, glucosilación, sulfatación, etc. Estas modifica-
manecen en él. ciones tienen lugar dentro de vesículas, las que por
• Proteínas de síntesis citosólica, pero con destino fi- transporte axoplasmático son transferidas hacia la
nal mitocondrial, nuclear o peroxisómico. membrana celular.
• Proteínas que se sintetizan en asociación con mem-
branas y se distribuyen por medio de vesículas en Puede así concluirse que las proteínas de membrana
distintas organelas. y las destinadas a la secreción son modificadas de ma-
nera significativa luego de su síntesis, a diferencia de
Las proteínas citoplasmáticas o citosólicas consti- lo que ocurre con las proteínas citosólicas. Los pro-
tuyen la fracción más importante y comprenden: a) ductos secretorios son sintetizados como parte de lar-
elementos fibrilares del citoesqueleto (neurofilamen- gas cadenas polipeptídicas, que sufren luego sucesivos
tos, tubulina y actina y proteínas asociadas que, en procesos de hidrólisis proteolítica.
conjunto, representan el 20% de las proteínas neuro- Los mecanismos de transferencia de las vesículas
nales), b) enzimas del metabolismo intermedio. Son desde el retículo endoplasmático al Golgi, y de allí a
proteínas sintetizadas en los polisomas libres y produ- los sitios de inserción en la membrana o de secreción,
cidas en su forma final, con muy poco procesado pos- son complejos. En las neuronas, las proteínas de mem-
terior y c) proteínas con destino mitocondrial, nuclear brana y de secreción son vehiculizadas a sus sitios fi-
o peroxisómico que también se sintetizan en poliso- nales por una de dos vías diferentes: a) en la vía cons-
mas libres, con inserción posterior en el sitio de desti- titutiva, las vesículas se mueven continuamente para
no (transferencia postraduccional). renovar el plasmalema, llevando nuevos constituyen-
Las proteínas de membrana y secretorias resultan tes y reciclando los viejos a través de los endosomas.
de la acción RNA mensajeros que forman polisomas Luego de ser recuperados del plasmalema, los endoso-
asociados con el retículo endoplasmático rugoso. La mas entran en los lisosomas para ser degradados, o son
sustancia de Nissl basófila, típica de las neuronas, es reciclados para reaparecer en la membrana plasmática
el resultado de la tinción de este RNA mensajero. La y b) en la vía regulada, las vesículas secretorias o si-
cadena peptídica comienza a sintetizarse por el N-ter- nápticas se fusionan con la membrana celular sólo en
minal, y existe una secuencia llamada péptido señal, el momento de la secreción que, como veremos, es de-
relativamente hidrófoba, que no permanece en la pro- pendiente del Ca2+ (fig. 1-23).
teína madura. El péptido señal tiene varias funciones.
Por un lado, le permite al polisoma unirse a la super- Cada sinapsis tiene un conjunto
ficie citoplasmática de la membrana del retículo endo- de receptores, canales y moléculas
plasmático. Asimismo, detiene la traducción del RNA
apropiadas para los neurotransmisores
mensajero. Por último, se libera péptido señal y la tra-
ducción recomienza. participantes
Según el destino final de la proteína, el péptido na-
ciente: Una cuestión clave en la biología de las neuronas es
comprender cómo los componentes celulares son diri-
• Se incorpora a porciones de la membrana del retícu- gidos a distancia desde el núcleo celular a muy distin-
Neurociencia aplicada ©2007. Editorial Médica Panamericana
26 Parte I - Los componentes del sistema nervioso y su comunicación
te a la proteína citoesquelética MAP-2 (microtubule-
associated protein; véase más adelante), y el que codi-
fica la síntesis de la subunidad alfa de la proteincinasa
dependiente de calmodulina. En menor proporción, en
las espinas dendríticas se encuentran ARNm corres-
pondientes a otros componentes del citoesqueleto. Los
ARNm mencionados se transportan asociados con los
componentes del citoesqueleto, por transporte axo-
plasmático lento.
En forma semejante a lo que ocurre en las neuronas,
se produce la síntesis de proteínas en regiones alejadas
del núcleo en células gliales. Por ejemplo, en los oli-
godendrocitos y en las células de Schwann, la proteí-
na básica de la mielina es sintetizada en los procesos
celulares (donde se encuentran los ARNm correspon-
dientes), mientras que los proteolípidos se sintetizan
perinuclearmente.
La función apropiada del sistema nervioso
depende del rápido y eficiente flujo de información
entre las neuronas y sus efectores, producido a
través de las sinapsis.
Fig. 1-23. Ciclo de vida de las vesículas sinápticas. Se sinteti- Si bien la morfología de la sinapsis se ha estudiado
zan, se ensamblan y se exportan desde el aparato de Golgi, durante mucho tiempo, sólo recientemente se ha obte-
transportándose por transporte axonal rápido hacia la sinapsis. nido información sobre las señales moleculares res-
Luego de la exocitosis y el reciclado retornan al cuerpo celular ponsables de la organización de estas estructuras.
por transporte retrógrado, donde se digieren en los lisosomas. La concentración selectiva de receptores es una de las
(Modificado de Kandel y col, 2000.)
propiedades típicas de la sinapsis. Los estudios más de-
tallados se han efectuado sobre el receptor nicotínico de
tos sitios del árbol dendrítico o del axón. Veremos más la placa muscular (véase cap. 2). En la sinapsis, la den-
adelante (cap. 3) que la función sináptica es el resulta- sidad de receptores es de unas 10.000 moléculas/mm2,
do de una particular combinación de proteínas (recep- mientras que fuera de la placa motora la densidad es
tores, canales iónicos, moléculas de adhesión y unas 1.000 veces menor. La principal molécula respon-
sistemas de segundos mensajeros), que determinan la sable de esta concentración es una proteína de 200 kDa
respuesta postsináptica al transmisor liberado en dicha producida por las motoneuronas y que se asocia con la
sinapsis. membrana postsináptica, llamada agrina. Esta proteína
tiene homologías con otros factores de crecimiento, co-
Por lo tanto, una neurona central, que recibe en mo el factor de crecimiento epidérmico.
promedio 104 sinapsis, debe construir 104 mi-
croambientes sinápticos que sean adecuados para El transporte axoplasmático es una
las variadas señales recibidas. adaptación funcional a la polaridad
Hasta hace poco se pensaba que estos microambien- extrema de las neuronas
tes se obtenían mediante los procesos de exportación
de proteínas desde el pericarion. Sin embargo, se ha Las neuronas son células secretorias. Como las célu-
identificado un segundo mecanismo dado por ARN las endocrinas, en las cuales los gránulos de secreción
mensajeros que se transfieren desde el núcleo neuro- se ensamblan en el aparato de Golgi, las neuronas pre-
nal a sitios sinápticos específicos para facilitar la sín- sentan vesículas de almacenamiento del transmisor
tesis local de proteínas. Ésta es la razón de que se en- (vesículas sinápticas), también formadas en el sistema
cuentren polirribosomas en dendritas, inmediatamente neuronal de membranas internas. A diferencia de las
por debajo de los sitios postsinápticos. Dos tipos de células glandulares, la extrema polarización de la neu-
ARNm predominan en las dendritas, el correspondien- rona hace que en muchos casos la distancia entre el
Neurociencia aplicada ©2007. Editorial Médica Panamericana
Biología de las células nerviosas 27
cuerpo celular y las terminales sinápticas sea conside- ronas presinápticas. Es de interés que tanto la activi-
rable. Líneas arriba hemos mencionado el ejemplo de dad eléctrica normal como las crisis convulsivas repe-
una motoneurona lumbar, con un axón varios órdenes tidas modifican la anatomía y la excitabilidad de las
de magnitud más largo que el diámetro del pericarion. redes neurales y la expresión de los genes que codifi-
Cobra así extrema importancia el tráfico de sustancias can la síntesis de neurotrofinas. Es probable que estos
entre el soma y las terminales o dendritas, denomina- mecanismos sean de importancia en procesos norma-
do transporte axoplasmático. les (p. ej., sueño, cap. 15; aprendizaje, cap. 16) y pa-
Existen dos tipos de transporte axoplasmático: tológicos (epilepsia, cap. 15).
Por transporte axoplasmático retrógrado, penetran el
• Anterógrado. SNC virus neurotrópicos como los agentes del herpes,
• Retrógrado. de la rabia y de la poliomielitis, así como toxinas (to-
xina tetánica).
Dentro del transporte axoplasmático anterógrado se El transporte axoplasmático anterógrado lento
distinguen los siguientes subgrupos: a) rápido y b) presenta dos componentes: a) velocidad de 0,5-3 mm/día
lento. y b) velocidad de 4-6 mm/día. A través del transpor-
En esencia, todas las organelas celulares que contie- te axoplasmático anterógrado lento viajan compo-
nen membranas se exportan desde el cuerpo celular nentes citosólicos (elementos del citoesqueleto y
por un proceso de transporte axoplasmático anteró- proteínas solubles). El subtipo más lento comprende
grado rápido, de velocidad promedio de 400 mm/día. las proteínas que forman los neurofilamentos y las
Los principales componentes transportados por que constituyen los microtúbulos (tubulina alfa y be-
este proceso son las vesículas sinápticas y las mito- ta y proteínas asociadas, como las MAP). El subtipo
condrias. más rápido de transporte axoplasmático anterógrado
Durante la exocitosis en las terminales neurales, las lento involucra a la actina (la cual al polimerizarse da
vesículas sinápticas se reciclan varias veces y la mem- origen a los microfilamentos) y a la clatrina (proteí-
brana celular es renovada constantemente por nuevos na que recubre vesículas en reciclado en el extremo
componentes que arriban desde el soma neuronal. A secretorio); la calmodulina también se desplaza en
fin de mantener un equilibrio entre los nuevos compo- este componente.
nentes de membrana que llegan y los que se reciclan Como puede apreciarse, los tres componentes prin-
en la terminal, estos últimos retornan al cuerpo celular cipales del citoesqueleto, microtúbulos, neurofilamen-
para su degradación o posterior reutilización. La velo- tos y microfilamentos, son transportados a través del
cidad de tal transporte axoplasmático retrógrado es axón y las dendritas por transporte axoplasmático an-
de unos 200 mm/día. terógrado lento.
Además de la función de reciclado de vesículas y de La forma de estudio de los distintos tipos de transpor-
la membrana celular, el transporte axoplasmático re- te axoplasmático consiste en la inyección de precursores
trógrado es utilizado para transferir al soma señales radiactivos (p. ej., aminoácidos) o de micropartículas ra-
producidas en elementos celulares postsinápticos, co- diactivas en las cercanías del soma neuronal y el segui-
mo por ejemplo, el factor de crecimiento neural. Es- miento de las moléculas marcadas a lo largo del axón.
te factor estimula el crecimiento de grupos neuronales Mediante este procedimiento se ha establecido que el
durante el desarrollo embriológico del SNC y tiene transporte axoplasmático anterógrado rápido es: a) de-
una posible aplicación en la recuperación del tejido pendiente de la fosforilación oxidativa, b) no es modifi-
neural adulto ante degeneraciones seniles o luego de la cado por inhibidores de la síntesis de proteínas, c) se
lesión. Pertenece a una familia más amplia de molécu- observa aun en axones desconectados del soma. Este
las tróficas neurales, llamadas neurotrofinas, que ac- transporte rápido está basado en los microtúbulos, que
túan sobre receptores vinculados a tirosincinasa y proveen una “vía” estacionaria sobre las cuales se mue-
constituyen señales de recuperación celular que impi- ven las organelas en forma saltatoria.
den la entrada de la célula en el proceso de apoptosis. El transporte axoplasmático anterógrado rápido de-
Las neurotrofinas de mayor importancia son el factor pende de varios de los filamentos que constituyen el
de crecimiento neural, la neurotrofina 3, la neurotrofi- citoesqueleto, es decir, la actina, la miosina y los mi-
na 4/5 y el factor neurotrófico cerebral (brain-deri- crotúbulos. Los microtúbulos proveen un “riel” sobre
ved neurotrophic factor, BDNF). el cual se mueven las partículas, y la traslocación, que
Todos pueden producirse en la postsinapsis como es dependiente de la energía, sería por deslizamiento
consecuencia de la actividad neural y son transporta- de filamentos de actina y miosina, en forma semejan-
dos por transporte axoplasmático retrógrado a las neu- te al proceso de contracción muscular (véase cap. 7).
Neurociencia aplicada ©2007. Editorial Médica Panamericana
28 Parte I - Los componentes del sistema nervioso y su comunicación
Son de importancia para definir la direccionalidad del
transporte axoplasmático anterógrado rápido y del re-
trógrado. Su longitud máxima en las dendritas o en el
axón es de unos 0,1 mm, no recorren toda la extensión
intracelular y no se continúan con microtúbulos del
cuerpo celular. Diversas proteínas asociadas (MAP-1,
MAP-2, tau) regulan la estabilidad de los microtúbu-
los y promueven su polimerización.
Los neurofilamentos, de 10 nm de diámetro, son
los elementos fibrilares más abundantes en los axo-
nes (10:1 en relación con los microtúbulos) y cons-
tituyen la base del citoesqueleto. Se denominan neu-
rofibrillas a los haces de neurofilamentos visibles al
microscopio óptico. Pertenecen, junto a los llama-
dos “filamentos intermedios” de otros tipos celula-
res, a la familia de proteínas de las citoqueratinas,
Fig. 1-24. Una MAP (proteína asociada con los microtúbulos),
que además comprende a la proteína fibrilar glial, a
la cinesina, de actividad ATPasa, está directamente vinculada
con el transporte axoplasmático anterógrado rápido. En pre- la desmina y a la queratina. Están totalmente
sencia de ATP, produce la fuerza necesaria para el desplaza- polimerizados en condiciones fisiológicas. En la en-
miento de las organelas. fermedad de Alzheimer se degeneran en forma ca-
racterística (los llamados tangles u ovillos de neuro-
filamentos). Una MAP (tau), fosforilada anormal-
mente, es responsable de este fenómeno.
Como hemos mencionado, los microtúbulos se com- Los microfilamentos, de 3-5 nm de diámetro, son
ponen de tubulina y proteínas asociadas (MAP). Una polímeros de actina en doble hélice. Su constitución es
de estas proteínas, la cinesina, de actividad ATPasa, semejante a la de la actina de otros grupos celulares.
está directamente vinculada con el transporte axoplas- En muchos casos, los microfilamentos se fijan a la
mático anterógrado rápido, y produce, en presencia de membrana celular a través de proteínas asociadas, co-
ATP, la fuerza necesaria para el desplazamiento de las mo la espectrina neuronal (o fodrina), la anquirina, la
organelas (fig. 1-24). Otra proteína de características vinculina y la talina. La mayoría de la actina neuronal
semejantes, la dineína, es la responsable del transpor- está asociada con la membrana celular; en las dendri-
te axoplasmático retrógrado. tas corticales se encuentra principalmente en las
Los elementos fibrilares del citoesqueleto neuronal espinas dendríticas, sitio de máxima abundancia de si-
se mueven por transporte axoplasmático lento. Estas napsis.
proteínas determinan la forma neuronal; presentan Los microfilamentos también pueden interaccionar
cambios de importancia en el envejecimiento normal y con proteínas de la matriz extracelular, como la la-
patológico (enfermedad de Alzheimer; cap. 16). minina o la fibronectina, asociándose con proteínas
que atraviesan la membrana, las integrinas. Estas
Las familias de proteínas fibrilares proteínas de superficie facilitan la adhesión y el
del citoesqueleto neuronal son tres reconocimiento celular y se unen a diversos compo-
nentes de la matriz extracelular, como la fibronecti-
Los principales elementos fibrilares del citoesquele- na, el colágeno o la laminina. Las integrinas se con-
to axonal son: sideran receptores para señales de la matriz extrace-
lular que afectan a la función celular. Su vía de
• Microtúbulos. segundo mensajero es la activación de la tirosincina-
• Neurofilamentos. sa (véase cap. 3).
• Microfilamentos (fig. 1-25). Los distintos componentes fibrilares del citoesquele-
to, en su conjunto, se hallan en estado dinámico, alar-
En cada caso se presentan también proteínas asocia- gándose o acortándose en forma continua. Por ejem-
das. plo, el 50% de la actina presente está en forma despo-
Los microtúbulos, compuestos por 13 protofila- limerizada; su polimerización se regula momento a
mentos de tubulina alfa y beta, tienen un diámetro de momento por complejos mecanismos intracelulares,
unos 25 nm, y están orientados longitudinalmente. aún no totalmente elucidados.
Neurociencia aplicada ©2007. Editorial Médica Panamericana
Biología de las células nerviosas 29
Fig. 1-25. Componentes del citoesqueleto neuronal.
BIBLIOGRAFÍA RECOMENDADA the brain matters for PET. Trends Neurosci 2005;28:117-9.
Ben Ari Y, Spitzer NC. Nature and nurture in brain develop-
Abbott NJ, Ronnback L, Hansson E. Astrocyte-endothelial in- ment. Trends Neurosci 2004;27:361.
teractions at the blood-brain barrier. Nat Rev Neurosci Benn SC, Woolf CJ. Adult neuron survival strategies– slam-
2006;7:41-53. ming on the brakes. Nat Rev Neurosci 2004;5:686-700.
Ackley BD, Jin Y. Genetic analysis of synaptic target recogni- Bennett MV, Contreras JE, Bukauskas FF, Saez JC. New roles
tion and assembly. Trends Neurosci 2004;27:540-7. for astrocytes: Gap junction hemichannels have something
Aidley DJ. The Physiology of Excitable Cells. London: Cam- to communicate. Trends Neurosci 2003;26:610-7.
bridge University Press; 1983. Chao MV. Neurotrophins and their receptors: A convergence
Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. point for many signalling pathways. Nat Rev Neurosci
Biología Molecular de la Célula. 4ª ed. Madrid: Omega; 2003;4:299-309.
2002. Chotard C, Salecker I. Neurons and glia: Team players in axon
Altenberg GA. The engine of ABC proteins. News Physiol Sci guidance. Trends Neurosci 2004;27:655-61.
2003;18:191-5. Chuckowree JA, Dickson TC, Vickers JC. Intrinsic regenerati-
Amiry-Moghaddam M, Ottersen OP. The molecular basis of ve ability of mature CNS neurons. Neuroscientist
water transport in the brain. Nat Rev Neurosci 2003;4:991- 2004;10:280-5.
1001. Conti L, Cattaneo E. Controlling neural stem cell division wit-
Anderson CM, Nedergaard M. Astrocyte-mediated control of hin the adult subventricular zone: An Appealing job. Trends
cerebral microcirculation. Trends Neurosci 2003;26:340-4. Neurosci 2005;28:57-9.
Andrews ZB, Diano S, Horvath TL. Mitochondrial uncoupling Crespo-Santiago D. La matriz extracelular del sistema nervio-
proteins in the CNS: In support of function and survival. Nat so central: Proteoglicanos del tipo condroitín sulfato y repa-
Rev Neurosci 2005;6:829-40. ración neural. Rev Neurol 2004;38:843-51.
Araujo SJ, Tear G. Axon guidance mechanisms and molecules: de Castro F. Chemotropic molecules: Guides for axonal path-
Lessons from invertebrates. Nat Rev Neurosci 2003;4:910-22. finding and cell migration during CNS development. News
Bareyre FM, Schwab ME. Inflammation, degeneration and re- Physiol Sci 2003;18:130-6.
generation in the injured spinal cord: insights from DNA Dean C, Dresbach T. Neuroligins and neurexins: Linking cell
microarrays. Trends Neurosci 2003;26:555-63. adhesion, synapse formation and cognitive function. Trends
Barros LF, Porras OH, Bittner CX. Why glucose transport in Neurosci 2006;29:21-9.
Neurociencia aplicada ©2007. Editorial Médica Panamericana
30 Parte I - Los componentes del sistema nervioso y su comunicación
Dent EW, Tang F, Kalil K. Axon guidance by growth cones and notype: What targets tell the cell bodies. Trends Neurosci
branches: Common cytoskeletal and signaling mechanisms. 2004;27:482-8.
Neuroscientist 2003;9:343-53. Hirokawa N, Takemura R. Molecular motors and mechanisms
Dillon C, Goda Y. The actin cytoskeleton: Integrating form and of directional transport in neurons. Nat Rev Neurosci
function at the synapse. Annu Rev Neurosci 2005;28:25-55. 2005;6:201-14.
Dityatev A, Schachner M. Extracellular matrix molecules and Horner PJ, Palmer TD. New roles for astrocytes: The nightlife
synaptic plasticity. Nat Rev Neurosci 2003;4:456-68. of an “astrocyte”. La vida loca! Trends Neurosci 2003;26:
Dorta-Contreras AJ, Reiber H. Teoría de la difusión molecular /flu- 597-603.
jo de líquido cefalorraquídeo. Rev Neurol 2004;39:564-9. Howard A, Tamas G, Soltesz I. Lighting the chandelier: New
Ebihara L. New roles for connexons. News Physiol Sci vistas for axo-axonic cells. Trends Neurosci 2005;28:310-6.
2003;18:100-3. Iadecola C. Neurovascular regulation in the normal brain and in
Ehlers MD. Ubiquitin and synaptic dysfunction: Ataxic mice Alzheimer’s disease. Nat Rev Neurosci 2004;5:347-60.
highlight new common themes in neurological disease. Innocenti GM, Price DJ. Exuberance in the development of
Trends Neurosci 2003;26:4-7. cortical networks. Nat Rev Neurosci 2005;6:955-65.
Emsley JG, Arlotta P, Macklis JD. Star-cross’d neurons: Astro- Jarjour AA, Kennedy TE. Oligodendrocyte precursors on the
glial effects on neural repair in the adult mammalian CNS. move: Mechanisms directing migration. Neuroscientist
Trends Neurosci 2004;27:238-40. 2004;10:99-105.
Esteban FJ, Horcajadas A, El Rubaidi O, Luque-Barona R, Iba- Jessen KR, Mirsky R. The origin and development of glial cells
ñez G, Garcáa-Carriazo A, Segovia M, Moral-Leal ML. in peripheral nerves. Nat Rev Neurosci 2005;6:671-82.
Óxido nítrico en astrocitos malignizados. Rev Neurol Jockusch BM, Huttelmaier S, Illenberger S. From the nucleus
2005;40:437-40. toward the cell periphery: A guided tour for mRNAs. News
Farber K, Kettenmann H. Physiology of microglial cells. Brain Physiol Sci 2003;18:7-11.
Res Brain Res Rev 2005;48:133-43. John GR, Lee SC, Brosnan CF. Cytokines: Powerful regulators
Ferguson KL, Slack RS. Growth factors: can they promote neu- of glial cell activation. Neuroscientist 2003;9:10-22.
rogenesis? Trends Neurosci 2003;26:283-5. Johnson MH. Functional brain development in humans. Nat
Ferrus A. El por qué de tantas sinapsis. Rev Neurol Rev Neurosci 2001;2:475-83.
2002;35:661-7. Kalb R. The protean actions of neurotrophins and their recep-
Freeman MR, Doherty J. Glial cell biology in Drosophila and tors on the life and death of neurons. Trends Neurosci
vertebrates. Trends Neurosci 2006;29:82-90. 2005;28:5-11.
Ganong WF. Review of Medical Physiology 21a ed. New York: Kandel ER, Schwartz JH, Jessel TM. Principles of Neural
McGraw-Hill. 2003 Science. 4th ed. New York: McGraw Hill; 2000.
Gandhi CS, Isacoff EY. Shedding light on membrane proteins. Kettenmann H, Ransom BR. Neuroglia. 2nd ed. Oxford: Oxford
Trends Neurosci 2005;28:472-9. University Press; 2004.
Garcia-Ovejero D, Azcoitia I, Doncarlos LL, Melcangi RC, Kim S, Chiba A. Dendritic guidance. Trends Neurosci 2004;27:
García-Segura LM. Glia-neuron crosstalk in the neuropro- 194-202.
tective mechanisms of sex steroid hormones. Brain Res Kirik D, Bjorklund A. Modeling CNS neurodegeneration by
Brain Res Rev 2005;48:273-86. overexpression of disease-causing proteins using viral vec-
Gidday JM. Cerebral preconditioning and ischaemic tolerance. tors. Trends Neurosci 2003;26:386-92.
Nat Rev Neurosci 2006;7:437-48. Klann E, Dever TE. Biochemical mechanisms for translational
Goldman S. Glia as neural progenitor cells. Trends Neurosci regulation in synaptic plasticity. Nat Rev Neurosci 2004;5:
2003;26:590-6. 931-42.
Gomez TM, Zheng JQ. The molecular basis for calcium-depen- Kleene R, Schachner M. Glycans and neural cell interactions.
dent axon pathfinding. Nat Rev Neurosci 2006;7:115-25. Nat Rev Neurosci 2004;5:195-208.
González-Amaro R, Sánchez-Madrid F. Moléculas de adhesión Lai CH, Kuo KH, Leo JM. Critical role of actin in modulating
intercelular y factores quimiotácticos en la patogenia de la BBB permeability. Brain Res Brain Res Rev 2005;50:7-13.
esclerosis múltiple. Rev Neurol 2002;35:985-93. Lauritzen M. Reading vascular changes in brain imaging: Is
Gotz M. Glial cells generate neurons– master control within dendritic calcium the key? Nat Rev Neurosci 2005;6:77-85.
CNS regions: Developmental perspectives on neural stem Lledo PM, Alonso M, Grubb MS. Adult neurogenesis and func-
cells. Neuroscientist 2003;9:379-97. tional plasticity in neuronal circuits. Nat Rev Neurosci
Guan KL, Rao Y. Signalling mechanisms mediating neuronal res- 2006;7:179-93.
ponses to guidance cues. Nat Rev Neurosci 2003;4:941-56. Lo EH, Dalkara T, Moskowitz MA. Mechanisms, challenges and
Hagg T. Molecular regulation of adult CNS neurogenesis: An opportunities in stroke. Nat Rev Neurosci 2003;4:399-415.
integrated view. Trends Neurosci 2005;28:589-95. Loturco JJ, Bai J. The multipolar stage and disruptions in neu-
Hanani M. Satellite glial cells in sensory ganglia: From form to ronal migration. Trends Neurosci 2006.
function. Brain Res Brain Res Rev 2005;48:457-76. Lu B, Pang PT, Woo NH. The yin and yang of neurotrophin ac-
Hanz S, Fainzilber M. Integration of retrograde axonal and nu- tion. Nat Rev Neurosci 2005;6:603-14.
clear transport mechanisms in neurons: implications for the- Luo L, O’Leary DD. Axon retraction and degeneration in deve-
rapeutics. Neuroscientist 2004;10:404-8. lopment and disease. Annu Rev Neurosci 2005;28:127-56.
Hatten ME, Heintz N. Large-scale genomic approaches to brain Ming GL, Song H. Adult neurogenesis in the mammalian cen-
development and circuitry. Annu Rev Neurosci 2005;28:89- tral nervous system. Annu Rev Neurosci 2005;28:223-50.
108. Missler M. Synaptic cell adhesion goes functional. Trends Neu-
Haydon PG. Glia: Listening and talking to the synapse. Nat rosci 2003;26:176-8.
Rev Neurosci 2001;2:185-93. Nedergaard M, Ransom B, Goldman SA. New roles for as-
Hertz L, Zielke HR. Astrocytic control of glutamatergic acti- trocytes: Redefining the functional architecture of the brain.
vity: Astrocytes as stars of the show. Trends Neurosci Trends Neurosci 2003;26:523-30.
2004;27:735-43. Newman EA. New roles for astrocytes: Regulation of synaptic
Hippenmeyer S, Kramer I, Arber S. Control of neuronal phe- transmission. Trends Neurosci 2003;26:536-42.
Neurociencia aplicada ©2007. Editorial Médica Panamericana
Biología de las células nerviosas 31
Notterpek L. Neurotrophins in myelination: A new role for a Seifert G, Schilling K, Steinhauser C. Astrocyte dysfunction in
puzzling receptor. Trends Neurosci 2003;26:232-4. neurological disorders: A molecular perspective. Nat Rev
Overstreet LS. Quantal transmission: not just for neurons. Neurosci 2006;7:194-206.
Trends Neurosci 2005;28:59-62. Shea TB, Jung C, Pant HC. Does neurofilament phosphoryla-
Pan W, Kastin AJ. Why study transport of peptides and proteins tion regulate axonal transport? Trends Neurosci 2003;26:
at the neurovascular interface. Brain Res Brain Res Rev 397-400.
2004;46:32-43. Shepherd GM. The Synaptic Organization of the Brain. Ox-
Pascual Garvi JM, González LF, Prieto AR, Cerdan S, Roda ford: Oxford University Press; 2004.
JM. La barrera hematoencefálica. Desarrollo de una estruc- Silver J, Miller JH. Regeneration beyond the glial scar. Nat Rev
tura que sostiene la heterogeneidad funcional del sistema Neurosci 2004;5:146-56.
nervioso central. Rev Neurol 2004;38:565-81. Smith R. Moving molecules: mRNA trafficking in mammalian
Paxinos G, Mai JK. The Human Nervous System. 2nd ed. New oligodendrocytes and neurons. Neuroscientist 2004;10:495-
York: Elsevier-Academic Press; 2004. 500.
Pellerin L, Magistretti PJ. Neuroenergetics: Calling upon as- Squire L, Bloom FE, McConnell SK, Roberts JL, Spitzer NC,
trocytes to satisfy hungry neurons. Neuroscientist Zigmond MJ. Fundamental Neuroscience. 2nd ed. New York:
2004;10:53-62. Academic Press; 2003.
Peters A, Palay SL, Webster H. The Fine Structure of the Ner- Theis M, Sohl G, Eiberger J, Willecke K. Emerging complexi-
vous System. The Neurons and Supporting Cells. New York: ties in identity and function of glial connexins. Trends Neu-
Oxford University Press; 1991. rosci 2005;28:188-95.
Purves D. Invitación a la Neurociencia. Buenos Aires: Editorial Tresguerres JAF, Ariznavarreta Ruiz C, Cachofeiro Ramos V,
Médica Panamericana; 2001. Cardinali D P, Escrich Escriche E, Gil Loyzaga P, Lahera
Rakic P. Neurogenesis in adult primate neocortex: An evalua- Juliá V, Mora Teruel F, Romano Pardo M, Tamargo Menén-
tion of the evidence. Nat Rev Neurosci 2002;3:65-71. dez J. Fisiología Humana. 3ª ed. Madrid: McGraw Hill-In-
Ramón y Cajal S. Textura del Sistema Nervioso del Hombre y teramericana; 2005.
de los Vertebrados. Alicante: Gráficas Vidal Leuka; 1992. Volterra A, Meldolesi J. Astrocytes, from brain glue to commu-
Rao SD, Weiss JH. Excitotoxic and oxidative cross-talk bet- nication elements: The revolution continues. Nat Rev Neu-
ween motor neurons and glia in ALS pathogenesis. Trends rosci 2005;6:626-40.
Neurosci 2004;27:17-23. Waagepetersen HS, Sonnewald U, Schousboe A. Compartmen-
Richardson WD, Kessaris N, Pringle N. Oligodendrocyte wars. tation of glutamine, glutamate, and GABA metabolism in
Nat Rev Neurosci 2006;7:11-8. neurons and astrocytes: Functional implications. Neuros-
Sawa A, Pletnikov MV, Kamiya A. Neuron-glia interactions cientist 2003;9:398-403.
clarify genetic-environmental links in mental illness. Trends Waites CL, Craig AM, Garner CC. Mechanisms of vertebrate
Neurosci 2004;27:294-7. synaptogenesis. Annu Rev Neurosci 2005;28:251-74.
Schaller B. Physiology of cerebral venous blood flow: From Yuste R, Bonhoeffer T. Genesis of dendritic spines: Insights
experimental data in animals to normal function in humans. from ultrastructural and imaging studies. Nat Rev Neurosci
Brain Res Brain Res Rev 2004;46:243-60. 2004;5:24-34.
Neurociencia aplicada ©2007. Editorial Médica Panamericana
Vous aimerez peut-être aussi
- Freud y Concepto de RegresionDocument5 pagesFreud y Concepto de RegresionZulema De La Riva CastilloPas encore d'évaluation
- Gomel Transmision Generacional Familia y Subjetividad Cap 6Document11 pagesGomel Transmision Generacional Familia y Subjetividad Cap 6Claudia Zuschterman100% (1)
- Amor PuroDocument2 pagesAmor PuroCesar RiveroPas encore d'évaluation
- Otto Kernberg sobre el narcisismo patológicoDocument8 pagesOtto Kernberg sobre el narcisismo patológicoLismarPas encore d'évaluation
- Semejanza Entre El Micro y El MacrocosmoDocument16 pagesSemejanza Entre El Micro y El MacrocosmoSophia GomezPas encore d'évaluation
- La Gestión de Las Tormentas Afectivas KernbergDocument37 pagesLa Gestión de Las Tormentas Afectivas KernbergLuis PerezPas encore d'évaluation
- Representación Fantasmática.Document4 pagesRepresentación Fantasmática.Ertp Itaf YoPas encore d'évaluation
- Erección Inventiva Jaques AndréDocument7 pagesErección Inventiva Jaques AndréjuanutoPas encore d'évaluation
- Conferencia 23Document2 pagesConferencia 23jimenavidelaPas encore d'évaluation
- De La Imago Materno A La Imagen PaternoDocument10 pagesDe La Imago Materno A La Imagen PaternoManuelPas encore d'évaluation
- Ejercicios Corporales para Liberar Bloqueos EnergéticosDocument2 pagesEjercicios Corporales para Liberar Bloqueos EnergéticosJonathan Alejandro ZambranoPas encore d'évaluation
- DinámicaTransferenciaDocument7 pagesDinámicaTransferenciaTania CardonaPas encore d'évaluation
- Multi FamiliarDocument31 pagesMulti FamiliarHyjo SamanPas encore d'évaluation
- PUGET-BERENSTEIN - El Zócalo Inconsciente de La ParejaDocument15 pagesPUGET-BERENSTEIN - El Zócalo Inconsciente de La ParejaAlejandra PaniaguaPas encore d'évaluation
- Fisiologia - Sistema - Nervioso - PPT ExpoDocument45 pagesFisiologia - Sistema - Nervioso - PPT ExpoSalud Medicina Confianza0% (1)
- La neurona como unidad funcional del SNCDocument33 pagesLa neurona como unidad funcional del SNCMaximiliano LozaPas encore d'évaluation
- Fisio Nervio 1Document7 pagesFisio Nervio 1Anadella RavalliPas encore d'évaluation
- Bases N Trab #4Document8 pagesBases N Trab #4Caro GePas encore d'évaluation
- Resumen Neuro B 2023 + Guía de TP 1, 2 y 3Document359 pagesResumen Neuro B 2023 + Guía de TP 1, 2 y 3MARíA SOLEDAD COSTILLAPas encore d'évaluation
- Cuestionario Unidad IIIDocument3 pagesCuestionario Unidad IIIJerali SaveryPas encore d'évaluation
- Introducción A La NeuropsicologiaDocument48 pagesIntroducción A La NeuropsicologiaAndrea SantiagoPas encore d'évaluation
- NeuronasDocument37 pagesNeuronasCarlangas Cortez ⃝⃒⃤Pas encore d'évaluation
- Apunte 1 UltraestructuraDocument7 pagesApunte 1 UltraestructuraRafahela Antonia Vergara RiveraPas encore d'évaluation
- 5991-BT-02-18-Fisiologà A Neuronal - Potencial Accià N e Impulso Nervioso - 7%Document20 pages5991-BT-02-18-Fisiologà A Neuronal - Potencial Accià N e Impulso Nervioso - 7%Luz Tapia VillarroelPas encore d'évaluation
- Resumen de NeuroanatomíaDocument49 pagesResumen de Neuroanatomíaadrian novaPas encore d'évaluation
- FISIOLOGIA DEL Sistema NerviosoDocument20 pagesFISIOLOGIA DEL Sistema NerviosoAndrea Agresti100% (1)
- Tejido nerviosoDocument8 pagesTejido nerviosoDubhé PeredaPas encore d'évaluation
- Clase 1 Desarrollo, Maduracioìn y Rganizacioìn Del SNC y SNPDocument21 pagesClase 1 Desarrollo, Maduracioìn y Rganizacioìn Del SNC y SNPkhendra matusPas encore d'évaluation
- Introduccion Basica VV 0.202110 PDFDocument19 pagesIntroduccion Basica VV 0.202110 PDFJeannette del RosarioPas encore d'évaluation
- Tema 7-2Document10 pagesTema 7-2saaraafdezz05Pas encore d'évaluation
- Introducción A La Neurociencia - BrainwareDocument51 pagesIntroducción A La Neurociencia - Brainwaremateo pianaPas encore d'évaluation
- Guía de Aprendizaje NeuronaDocument4 pagesGuía de Aprendizaje Neuronatercerom2009100% (2)
- NeuronasDocument37 pagesNeuronasFlavio lescart100% (1)
- Anatomia e Histologia HumanaDocument33 pagesAnatomia e Histologia HumanaGuillermo TorrePas encore d'évaluation
- NeuroDocument14 pagesNeuroMicaelaPas encore d'évaluation
- Fisiologia de La Neurona IDocument7 pagesFisiologia de La Neurona IJaviera AriasPas encore d'évaluation
- F NeuronasDocument5 pagesF NeuronasSpreen ClipsPas encore d'évaluation
- Introducción Al Estudio Del Sistema NerviosoDocument20 pagesIntroducción Al Estudio Del Sistema Nerviosoabraham aucapiñaPas encore d'évaluation
- Taller Guía Estudio Sistema Nervioso Catedra Morfofisiología Bacteriología y L.C. Universidad de SantanderDocument16 pagesTaller Guía Estudio Sistema Nervioso Catedra Morfofisiología Bacteriología y L.C. Universidad de Santandermayerly ariasPas encore d'évaluation
- NeuronaDocument8 pagesNeuronaReyna RodriguezPas encore d'évaluation
- Histología Del Sistema Nervioso y Liberación QuímicaDocument33 pagesHistología Del Sistema Nervioso y Liberación QuímicaDiego CeliPas encore d'évaluation
- Practica Tejido Nervioso Llena ARREGLADADocument5 pagesPractica Tejido Nervioso Llena ARREGLADABioqui MicaPas encore d'évaluation
- HistologiaDocument11 pagesHistologiaLISETHPas encore d'évaluation
- Introducción a la neurona: Estructura, función y plasticidadDocument19 pagesIntroducción a la neurona: Estructura, función y plasticidadRoxana Montenegro100% (1)
- Anatomía de La NeuronaDocument7 pagesAnatomía de La NeuronaClaudita G. Domínguez PradoPas encore d'évaluation
- El Sistema Nervioso 809Document26 pagesEl Sistema Nervioso 809Luis Alejandro Ebratt MendozaPas encore d'évaluation
- Histologia, Medula EspinalDocument47 pagesHistologia, Medula EspinalJuana PatanaPas encore d'évaluation
- Investigacion Sistema NerviosoDocument23 pagesInvestigacion Sistema NerviosoValdo CruzPas encore d'évaluation
- Sistema NerviosoDocument11 pagesSistema NerviosoSaúl Ramírez IñigoPas encore d'évaluation
- Tejido Nervioso ApunteDocument39 pagesTejido Nervioso ApunteFacu CobosPas encore d'évaluation
- Neurona estructura función 40Document15 pagesNeurona estructura función 40Nico Ignacio VeraPas encore d'évaluation
- Anatomia y Fisiología Semana 6 - 2021 IiiDocument8 pagesAnatomia y Fisiología Semana 6 - 2021 IiiDavid LlontopPas encore d'évaluation
- HISTOLOGÍA VETERINARIA - Tejido NerviosoDocument19 pagesHISTOLOGÍA VETERINARIA - Tejido NerviosoMarielita Cepeda100% (1)
- #1 La NeuronaDocument11 pages#1 La NeuronaMonserrat Olivo TalaveraPas encore d'évaluation
- Tejido Nerviso 1, 2do AñoDocument10 pagesTejido Nerviso 1, 2do AñoMercedes SánchezPas encore d'évaluation
- Neurona Unidad Funcional y Estructural IDocument10 pagesNeurona Unidad Funcional y Estructural ICarlos TolentinoPas encore d'évaluation
- Sistema nervioso y neuroanatomíaDocument31 pagesSistema nervioso y neuroanatomíaMaritza GonzalezPas encore d'évaluation
- Tarea 7.1 Tejido Nervioso HISTO 1Document7 pagesTarea 7.1 Tejido Nervioso HISTO 1Elizabeth EstevezPas encore d'évaluation
- Tejido Nervioso AnimalDocument27 pagesTejido Nervioso AnimalCarmen Luzmila Carlos de la CruzPas encore d'évaluation
- GlosarioDocument9 pagesGlosarioEleazar PerezPas encore d'évaluation
- Tesis Sobre Lesiones Cerebrales PDFDocument158 pagesTesis Sobre Lesiones Cerebrales PDFLuis LuisPas encore d'évaluation
- Quimica y Fisiología Del Sistema NerviosoDocument104 pagesQuimica y Fisiología Del Sistema NerviosoLuciano MirianiPas encore d'évaluation
- Inteligencia y MielinaDocument4 pagesInteligencia y MielinaFrarodri100% (1)
- Clasificacion de Fibras NerviosasDocument49 pagesClasificacion de Fibras NerviosasVictor Black100% (1)
- Sistema Nervioso EmbriologiaDocument32 pagesSistema Nervioso Embriologiaramon ReyesPas encore d'évaluation
- 9277 9524 1 PBDocument28 pages9277 9524 1 PBKarito YancuntaPas encore d'évaluation
- SNCDocument14 pagesSNCYOCELINPas encore d'évaluation
- Teoria Informacional de LapersonalidadDocument78 pagesTeoria Informacional de LapersonalidadGerardo LuisPas encore d'évaluation
- Preguntas de Tejido Sanguíneo, Nervioso y MuscularDocument2 pagesPreguntas de Tejido Sanguíneo, Nervioso y MuscularLenin Lopez100% (2)
- Miastenia GravisDocument22 pagesMiastenia GravisAngela DanielaPas encore d'évaluation
- Mitos y literatura prehispánicaDocument13 pagesMitos y literatura prehispánicaDianaAfricano100% (1)
- Relación entre el sistema nervioso y otros sistemas del organismoDocument19 pagesRelación entre el sistema nervioso y otros sistemas del organismovictoria sofiaPas encore d'évaluation
- Glosario de Neurociencias PsiDocument32 pagesGlosario de Neurociencias PsiFernanda BustamantePas encore d'évaluation
- Clases de Células GlialesDocument2 pagesClases de Células GlialesJhoselyn Zarait Castillo GonzalesPas encore d'évaluation
- 1.1 Somestesia. Potencial de Membrana. SinapsisDocument92 pages1.1 Somestesia. Potencial de Membrana. SinapsisFLAVIA GAZZELLA .Pas encore d'évaluation
- GLIOBLASTOSDocument27 pagesGLIOBLASTOSMarintalero100% (1)
- Tarea No.3 Sistema NerviosoDocument6 pagesTarea No.3 Sistema NerviosoDiego AlvaradoPas encore d'évaluation
- Preguntas Del Curso de EnfermeriaDocument51 pagesPreguntas Del Curso de EnfermeriaAromi SaRoPas encore d'évaluation
- Desarrollo humano 0-3 añosDocument6 pagesDesarrollo humano 0-3 añosDavid Reyes PradoPas encore d'évaluation
- Células Gliales y Neuronas BASES BIOLOGICASDocument19 pagesCélulas Gliales y Neuronas BASES BIOLOGICASAmérica CastilloPas encore d'évaluation
- Guia Histologia 2 TerminadaDocument30 pagesGuia Histologia 2 TerminadaEfrain MalpicaPas encore d'évaluation
- Enfermedades Autoinmunitarias 2018Document92 pagesEnfermedades Autoinmunitarias 2018Alex100% (1)
- Trastornos Por NeurotoxicidadDocument56 pagesTrastornos Por NeurotoxicidadMuy chevere RobotPas encore d'évaluation
- 8089-3815-Solucionario Control U1 III BioDocument7 pages8089-3815-Solucionario Control U1 III BioFranPas encore d'évaluation
- Neurotransmisores y HomestatasisDocument8 pagesNeurotransmisores y HomestatasisAmaroPas encore d'évaluation
- Acido Lipoico y Neuropatia DiabticaDocument8 pagesAcido Lipoico y Neuropatia DiabticaDaniel GarzaPas encore d'évaluation
- Esclerosis Multiple TerminadoDocument16 pagesEsclerosis Multiple TerminadoRomina Callapani Montoya100% (1)
- Tumores benignos y malignos de la cavidad oralDocument21 pagesTumores benignos y malignos de la cavidad oralRenatito ValdiviezoPas encore d'évaluation
- Psicofisio 1.0 PDFDocument9 pagesPsicofisio 1.0 PDFAriel VillarroelPas encore d'évaluation
- Separata DPS 2023-1 Parte 1Document147 pagesSeparata DPS 2023-1 Parte 1Sebastian Jared Davila GonzalesPas encore d'évaluation