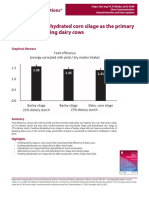
Vous aimerez peut-être aussi
- Diet System of Milking Cows With Sillage of Wet CornDocument9 pagesDiet System of Milking Cows With Sillage of Wet CornCk_psihPas encore d'évaluation
- Effect of Dietary Protein Source On Feed Intake, Growth, Pancreatic Enzyme and Jejunal Morphology Newly-Weaned PigletsDocument16 pagesEffect of Dietary Protein Source On Feed Intake, Growth, Pancreatic Enzyme and Jejunal Morphology Newly-Weaned PigletsSivik LudwyPas encore d'évaluation
- Dextrin SDocument5 pagesDextrin SRiezal BintanPas encore d'évaluation
- The Effects of Condensed Tannins From Desmodium Intortum and Calliandra Calothyrsus On Protein and Carbohydrate Digestion in Sheep and GoatsDocument19 pagesThe Effects of Condensed Tannins From Desmodium Intortum and Calliandra Calothyrsus On Protein and Carbohydrate Digestion in Sheep and GoatsMai elvi KhairaniPas encore d'évaluation
- Digestion of Starch and Sugars: E. Blas and T. GidenneDocument22 pagesDigestion of Starch and Sugars: E. Blas and T. GidenneAlejandro BoteroPas encore d'évaluation
- Dias Et Al., 2022Document10 pagesDias Et Al., 2022jumoro13Pas encore d'évaluation
- Diet Formulation Broilers Based Total Versus Amino Acids: DigestibleDocument7 pagesDiet Formulation Broilers Based Total Versus Amino Acids: Digestiblecelia cjanahuirePas encore d'évaluation
- Nutrition Reviews - February 1986 - Wurzburg - Nutritional Aspects and Safety of Modified Food StarchesDocument6 pagesNutrition Reviews - February 1986 - Wurzburg - Nutritional Aspects and Safety of Modified Food Starchesdiana salazarPas encore d'évaluation
- Swine Feed FomulationDocument6 pagesSwine Feed FomulationLok Raj JoshiPas encore d'évaluation
- Michigan State University, East LansingDocument5 pagesMichigan State University, East LansingElizabeth Alarcón HernándezPas encore d'évaluation
- Accepted Manuscript: Enzyme and Microbial TechnologyDocument41 pagesAccepted Manuscript: Enzyme and Microbial TechnologyTidar MelianiPas encore d'évaluation
- First Page PDFDocument1 pageFirst Page PDFladywhite820Pas encore d'évaluation
- 2017 Digestible Indispensable Amino Acid ScoreDocument5 pages2017 Digestible Indispensable Amino Acid ScoreROSSALINA BINTI ABDULLAH / UPMPas encore d'évaluation
- Starch ST Rke - 2018 - McCleary - Measurement of Starch Critical Evaluation of Current MethodologyDocument13 pagesStarch ST Rke - 2018 - McCleary - Measurement of Starch Critical Evaluation of Current MethodologySaha EtaPas encore d'évaluation
- 2.02 Energy and Ecosystem Lab: Soybean Grasshopp Er Chicken FarmerDocument3 pages2.02 Energy and Ecosystem Lab: Soybean Grasshopp Er Chicken FarmerAlexis Williams100% (2)
- Molecular Characterisation of Carbohydrate Digestion and Absorption in Equine Small IntestineDocument10 pagesMolecular Characterisation of Carbohydrate Digestion and Absorption in Equine Small IntestineArthur HeviPas encore d'évaluation
- Erdman 1989Document4 pagesErdman 1989leo06031314Pas encore d'évaluation
- Agricultural Research Bulletin v032 b443Document27 pagesAgricultural Research Bulletin v032 b443Saroj Kumar RaiPas encore d'évaluation
- PNS PAES 413 2001 Agricultural Structures Biogas PlantDocument38 pagesPNS PAES 413 2001 Agricultural Structures Biogas PlantKrystian BonghanoyPas encore d'évaluation
- Gastrointestinal Morphology and Absorption of Monosaccharides in Conditioned To Different Types and Levels of Dietary FibreDocument13 pagesGastrointestinal Morphology and Absorption of Monosaccharides in Conditioned To Different Types and Levels of Dietary FibreayakashimajorinPas encore d'évaluation
- Pretreatment and Saccharification of Rice HuksDocument7 pagesPretreatment and Saccharification of Rice HuksHimpunan Mahasiswa Teknik KimiaPas encore d'évaluation
- Partial or Total Replacement of Fishmeal by Solvent-Extracted Cottonseed Meal in Diets For Juvenile Rainbow Trout (Oncorhynchus Mykiss)Document8 pagesPartial or Total Replacement of Fishmeal by Solvent-Extracted Cottonseed Meal in Diets For Juvenile Rainbow Trout (Oncorhynchus Mykiss)Cherry VelasquezPas encore d'évaluation
- From By-Product To Valuable Components: Efficient Enzymatic Conversion of Lactose in Whey Using Galactosidase From Streptococcus ThermophilusDocument9 pagesFrom By-Product To Valuable Components: Efficient Enzymatic Conversion of Lactose in Whey Using Galactosidase From Streptococcus ThermophilusAntonela PortaPas encore d'évaluation
- Starch 1Document17 pagesStarch 1Umut ErkılıçPas encore d'évaluation
- Molasses in Beef NutritionDocument57 pagesMolasses in Beef NutritionMinh PhamPas encore d'évaluation
- Energy Value of Oatmeal and The Digestibility and Absorption of Its Proteins Fats and CalciumDocument8 pagesEnergy Value of Oatmeal and The Digestibility and Absorption of Its Proteins Fats and CalciumChinmaya MishraPas encore d'évaluation
- Pea Protein IsolatesDocument23 pagesPea Protein IsolatesqixiadeyouxiangPas encore d'évaluation
- Feed Requirements of CamelDocument10 pagesFeed Requirements of CamelNauman AzeemiPas encore d'évaluation
- NotesDocument76 pagesNotesAdrianne Joy De CastroPas encore d'évaluation
- A McCleary 2014Document7 pagesA McCleary 2014Cristhian OrtizPas encore d'évaluation
- Crude Fiber Lab Report - Docx Updated FileDocument7 pagesCrude Fiber Lab Report - Docx Updated FileNicholas Boampong100% (2)
- Effect of Ammonia Concentration On Rumen Microbial Protein Production in VitroDocument10 pagesEffect of Ammonia Concentration On Rumen Microbial Protein Production in VitroHidayat Anjar AsmaraniPas encore d'évaluation
- The Digestion of Cellulose by SheepDocument5 pagesThe Digestion of Cellulose by Sheepnutrisionist06Pas encore d'évaluation
- Ty11 PDFDocument8 pagesTy11 PDFJUANPas encore d'évaluation
- Lukondeh 2005Document5 pagesLukondeh 2005AleLeonPas encore d'évaluation
- Properties and Applications of Starch-Converting Enzymes of The A-Amylase FamilyDocument19 pagesProperties and Applications of Starch-Converting Enzymes of The A-Amylase Familya1805352Pas encore d'évaluation
- Abstracts of Communications PDFDocument87 pagesAbstracts of Communications PDFMusabyassinPas encore d'évaluation
- Protein Degradation in RumenDocument12 pagesProtein Degradation in Rumenjerson stiven moreno blancoPas encore d'évaluation
- Effect of Maltose On Glucoamylase Formation by Aspergillus NigerDocument7 pagesEffect of Maltose On Glucoamylase Formation by Aspergillus NigerKukymovPas encore d'évaluation
- Determination of The Net Utilization of Proteins by A Shortened MethodDocument7 pagesDetermination of The Net Utilization of Proteins by A Shortened MethodSrujanKumarPas encore d'évaluation
- CoA y L-CarnitinaDocument5 pagesCoA y L-CarnitinaDiane FloresPas encore d'évaluation
- Int J of Food Sci Tech - October 1976 - YONG - Microbial Succession in Experimental Soy Sauce FermentationsDocument12 pagesInt J of Food Sci Tech - October 1976 - YONG - Microbial Succession in Experimental Soy Sauce FermentationsSantosh KumarPas encore d'évaluation
- Cassava SilageDocument8 pagesCassava SilageblechikotiPas encore d'évaluation
- Bi 9740067Document8 pagesBi 9740067Bachi VarmaPas encore d'évaluation
- Starch ST Rke - 2018 - McCleary - Measurement of Starch Critical Evaluation of Current MethodologyDocument13 pagesStarch ST Rke - 2018 - McCleary - Measurement of Starch Critical Evaluation of Current Methodologyqinhuixing3Pas encore d'évaluation
- Tebon Jagung 2Document5 pagesTebon Jagung 2Siti AlfinaPas encore d'évaluation
- Art. SULMA PDFDocument6 pagesArt. SULMA PDFSulma Salas DeldadoPas encore d'évaluation
- Crude Glycerol in The Diets of The Juveniles Ofamazon Catfish (Female Pseudoplatystoma Punctifer X Male Leiarius Marmoratus)Document16 pagesCrude Glycerol in The Diets of The Juveniles Ofamazon Catfish (Female Pseudoplatystoma Punctifer X Male Leiarius Marmoratus)IJEAB JournalPas encore d'évaluation
- Assignments On Ration Formulation For Dairy Cattle FeedingDocument12 pagesAssignments On Ration Formulation For Dairy Cattle FeedingJoseph NdunguPas encore d'évaluation
- 7 BufalosDocument108 pages7 BufalosTauseefAhmadPas encore d'évaluation
- Cover Page: Picture of Chamber With Cow (Michal?)Document4 pagesCover Page: Picture of Chamber With Cow (Michal?)Ioan MoldovanPas encore d'évaluation
- Ijiaar 667 Manuscript 195006 1Document16 pagesIjiaar 667 Manuscript 195006 1Teodora Popova UmutesiPas encore d'évaluation
- Note Extender Double-Mutant Rice Starch Granules Does Not Determine TheirDocument6 pagesNote Extender Double-Mutant Rice Starch Granules Does Not Determine TheirTiara Kurnia Khoerunnisa FapertaPas encore d'évaluation
- Upgrading of Animal Feed by Solid State Fermentation by Pleurotus Sajor-CajuDocument6 pagesUpgrading of Animal Feed by Solid State Fermentation by Pleurotus Sajor-CajuKareem LashinPas encore d'évaluation
- MesophilDocument4 pagesMesophilnurcincelikPas encore d'évaluation
- Comparison of Grass and Legume Silages For Milk Production. 2. in Vivo and in Sacco Evaluations of Rumen FunctionDocument10 pagesComparison of Grass and Legume Silages For Milk Production. 2. in Vivo and in Sacco Evaluations of Rumen FunctionIoan MoldovanPas encore d'évaluation
- Plphys v46 3 429Document6 pagesPlphys v46 3 429Christian Carl GimenaPas encore d'évaluation
- Dietary Rhubarb Rheum Rhaponticum Stalk Fibre Stimulates Cholesterol 7 Hydroxylase Gene Expression and Bile Acid Excretion in Cholesterol Fed c57bl 6j MiceDocument7 pagesDietary Rhubarb Rheum Rhaponticum Stalk Fibre Stimulates Cholesterol 7 Hydroxylase Gene Expression and Bile Acid Excretion in Cholesterol Fed c57bl 6j MiceRadita AgustiniPas encore d'évaluation
- Camel Feed RequirementDocument11 pagesCamel Feed RequirementabdirahmanPas encore d'évaluation
- Nutritional Management of Equine Diseases and Special CasesD'EverandNutritional Management of Equine Diseases and Special CasesBryan M. WaldridgePas encore d'évaluation
- EndoscopyNovianaetal2017JKH PDFDocument6 pagesEndoscopyNovianaetal2017JKH PDFJuni ClaudiaPas encore d'évaluation
- Smith2002 PDFDocument37 pagesSmith2002 PDFJuni ClaudiaPas encore d'évaluation
- Animal Models in Type 2 Diabetes Research: December 2015Document30 pagesAnimal Models in Type 2 Diabetes Research: December 2015Juni ClaudiaPas encore d'évaluation
- Canine Chronic Kidney Disease Current Diagnostics & Goals For Long-Term ManagementDocument7 pagesCanine Chronic Kidney Disease Current Diagnostics & Goals For Long-Term ManagementJuni ClaudiaPas encore d'évaluation
- Embryonic Stem Cell Models of Development: TutorialDocument10 pagesEmbryonic Stem Cell Models of Development: TutorialJuni ClaudiaPas encore d'évaluation
- Excess Parathyroid Hormone Adversely Affects Lipid Metabolism in Chronic Renal FailureDocument5 pagesExcess Parathyroid Hormone Adversely Affects Lipid Metabolism in Chronic Renal FailureJuni ClaudiaPas encore d'évaluation
- Jurnal CKDDocument7 pagesJurnal CKDJuni ClaudiaPas encore d'évaluation
- (EAAP Scientific Series) Lapierre, Hélène - Skomiał, Jacek - Energy and Protein Metabolism and Nutrition 2016-Wageningen Academic Publishers (2016)Document385 pages(EAAP Scientific Series) Lapierre, Hélène - Skomiał, Jacek - Energy and Protein Metabolism and Nutrition 2016-Wageningen Academic Publishers (2016)Juni ClaudiaPas encore d'évaluation
- JBC Hist Persp Lipids PDFDocument67 pagesJBC Hist Persp Lipids PDFJuni ClaudiaPas encore d'évaluation
- BabesiosisDocument20 pagesBabesiosisJuni ClaudiaPas encore d'évaluation
- Aniceta Palacio v. SudarioDocument2 pagesAniceta Palacio v. SudarioLorebeth EspañaPas encore d'évaluation
- Review On Compound Animal Feed Processing in Ethiopia: Condition, Challenges and OpportunitiesDocument5 pagesReview On Compound Animal Feed Processing in Ethiopia: Condition, Challenges and OpportunitiesFeyisa RetaPas encore d'évaluation
- Animal Husbandry Preamble: For Candidates in Ghana OnlyDocument23 pagesAnimal Husbandry Preamble: For Candidates in Ghana OnlyZainabPas encore d'évaluation
- The Potential of Ettawa Goat Manure and Urine Management To Support The Productive and Sustainable FarmingDocument5 pagesThe Potential of Ettawa Goat Manure and Urine Management To Support The Productive and Sustainable FarmingBagas Septya PradanaPas encore d'évaluation
- Group-23 V.L.D.A.Document3 pagesGroup-23 V.L.D.A.Sahil KunduPas encore d'évaluation
- How To Feed Your Chickens Without Grain - 20 Ways To Cut Your Costs 100% - Abundant PermacultureDocument33 pagesHow To Feed Your Chickens Without Grain - 20 Ways To Cut Your Costs 100% - Abundant PermacultureJhanese Bastes SarigumbaPas encore d'évaluation
- Hereumu Herald - Nov 2015Document6 pagesHereumu Herald - Nov 2015Anaru MillPas encore d'évaluation
- NDDB Maharashtra Dairy Diggest PDFDocument147 pagesNDDB Maharashtra Dairy Diggest PDFManoj Tribhuwan100% (1)
- A 0242 e 00Document252 pagesA 0242 e 00Vijay KumarPas encore d'évaluation
- Ian Christopher Scoones 1990 PHD ThesisDocument585 pagesIan Christopher Scoones 1990 PHD ThesisAnonymous xe5sgvKWCPas encore d'évaluation
- Milk Production of Hand Milked Dairy Cattle in Burkina FasoDocument86 pagesMilk Production of Hand Milked Dairy Cattle in Burkina Fasofranky100% (1)
- Streptomycin PenicillinDocument76 pagesStreptomycin PenicillinMuhammad ZeeshanPas encore d'évaluation
- 2052 Compounded Cattle FeedDocument16 pages2052 Compounded Cattle FeedRISHI FOOD TESTING LABPas encore d'évaluation
- Hannah Velten: Nimal SeriesDocument210 pagesHannah Velten: Nimal SeriesSivaraman VisvanathanPas encore d'évaluation
- WGS243 Assessment 6Document3 pagesWGS243 Assessment 6Diggie FeederPas encore d'évaluation
- Farm Model of Dairy Unit of 10 Crossbred CowsDocument3 pagesFarm Model of Dairy Unit of 10 Crossbred CowsKarthik RamakrishnanPas encore d'évaluation
- HDocument4 pagesHwisnuPas encore d'évaluation
- EF4e Uppint Progress 6 10aDocument9 pagesEF4e Uppint Progress 6 10aJanneth YanzaPas encore d'évaluation
- Dairy ProjectDocument14 pagesDairy ProjectAdnan Hossain100% (1)
- 2019 Barham Koondrook Show ScheduleDocument33 pages2019 Barham Koondrook Show ScheduleJulietPas encore d'évaluation
- 12 Golden Rules For Milking - DHPDocument3 pages12 Golden Rules For Milking - DHPdremerboyPas encore d'évaluation
- 98 Leigh Park History 1874 To 1904Document148 pages98 Leigh Park History 1874 To 1904Ralph CousinsPas encore d'évaluation
- Moot Problem Constitutional B.A.LL.B., B.B.A.LL.B. and LL.B.Document22 pagesMoot Problem Constitutional B.A.LL.B., B.B.A.LL.B. and LL.B.ishitaPas encore d'évaluation
- Geography-ECONOMIC GEOGRAPHY REVIEW Revised PDFDocument97 pagesGeography-ECONOMIC GEOGRAPHY REVIEW Revised PDFnarishaPas encore d'évaluation
- Computation of Ration For Farm Animals-037Document21 pagesComputation of Ration For Farm Animals-037lalit_shivhare33% (6)
- Beef Cattle Behavior and HandlingDocument7 pagesBeef Cattle Behavior and Handlingbinodtharu0014Pas encore d'évaluation
- Reproductive Technologies in Farm AnimalsDocument347 pagesReproductive Technologies in Farm AnimalsClecio Henrique Limeira100% (1)
- Hubungan Volume Ambing Dan Ukuran Puting Dengan Produksi Susu Sapi Perah Friesian Holstein Di PT. Naksatra Kejora, Kabupaten TemanggungDocument9 pagesHubungan Volume Ambing Dan Ukuran Puting Dengan Produksi Susu Sapi Perah Friesian Holstein Di PT. Naksatra Kejora, Kabupaten TemanggungTiara RahmayaniPas encore d'évaluation
- Commercial Dairy Goat FarmsDocument63 pagesCommercial Dairy Goat FarmsKlaudija LutovskaPas encore d'évaluation
- Pen Fattenning of Beef Cattle ExplainedDocument7 pagesPen Fattenning of Beef Cattle ExplainedimugabePas encore d'évaluation