Vous aimerez peut-être aussi
- EAD - 0 A 18 MesesDocument9 pagesEAD - 0 A 18 MesesEyder Alexander RodriguezPas encore d'évaluation
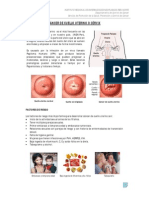
- CIANOSISDocument3 pagesCIANOSISEyder Alexander RodriguezPas encore d'évaluation
- EAD - 0 A 7 AñosDocument14 pagesEAD - 0 A 7 AñosEyder Alexander RodriguezPas encore d'évaluation
- GINA 2020 Traducción BásicaDocument154 pagesGINA 2020 Traducción BásicaEyder Alexander RodriguezPas encore d'évaluation
- Los 10 Mandamientos Del Médico InternoDocument8 pagesLos 10 Mandamientos Del Médico InternoEyder Alexander RodriguezPas encore d'évaluation
- Neisseria Gonorrhoeae PDFDocument2 pagesNeisseria Gonorrhoeae PDFEyder Alexander RodriguezPas encore d'évaluation
- Quiz Sobre Patología MamariaDocument4 pagesQuiz Sobre Patología MamariaEyder Alexander RodriguezPas encore d'évaluation
- Principios de Bioética Cambra F.JDocument9 pagesPrincipios de Bioética Cambra F.JEyder Alexander RodriguezPas encore d'évaluation
- BioéticaDocument5 pagesBioéticaEyder Alexander RodriguezPas encore d'évaluation
- Introducción.: Tema: ¿Por Qué No Nos Enfermamos Con Los Desechos Que Produce El Cuerpo?Document18 pagesIntroducción.: Tema: ¿Por Qué No Nos Enfermamos Con Los Desechos Que Produce El Cuerpo?Eyder Alexander RodriguezPas encore d'évaluation
- Diagnóstico Virológico Por LaboratorioDocument19 pagesDiagnóstico Virológico Por LaboratorioEyder Alexander RodriguezPas encore d'évaluation
- Programa Toxina RemovedDocument5 pagesPrograma Toxina RemovedDr. Felipe BauerPas encore d'évaluation
- ANESTESICOSDocument33 pagesANESTESICOSWilhem Roberth Soto FloresPas encore d'évaluation
- Sesiòn 3Document6 pagesSesiòn 3DIANA VALENTINA RIVAS FERNANDEZPas encore d'évaluation
- Enfoque Del Adulto Con Fiebre de Origen Desconocido - UpToDateDocument32 pagesEnfoque Del Adulto Con Fiebre de Origen Desconocido - UpToDateAlan Gustavo Arellano RamosPas encore d'évaluation
- Taller de Clasificacixn de Enfermedades e Intervenciones. CIE-10 y CIE-9MC. CEMECE. Instr. Rodolfo Alanxs FuentesDocument54 pagesTaller de Clasificacixn de Enfermedades e Intervenciones. CIE-10 y CIE-9MC. CEMECE. Instr. Rodolfo Alanxs FuentesGisela Camarillo MoralesPas encore d'évaluation
- Enfermería - Malla Curricula 2022Document1 pageEnfermería - Malla Curricula 2022Lidia Navarro100% (1)
- Mordeduras y Cuerpos ExtrañosDocument35 pagesMordeduras y Cuerpos ExtrañosGladysElizabethAlmarzaRamirezPas encore d'évaluation
- Transfusion F-FDocument6 pagesTransfusion F-FROsadelaCruzPas encore d'évaluation
- Ficha Deportiva JugadorDocument2 pagesFicha Deportiva JugadorGrettell Ivonne Báez garcia100% (2)
- Análisis de Psicología ComunitariaDocument3 pagesAnálisis de Psicología ComunitariaOscar BarrazaPas encore d'évaluation
- Landa EspinozaDocument34 pagesLanda EspinozaRudyMantariLauraPas encore d'évaluation
- Farmacologia Tarea 5 AntiparacitariosDocument11 pagesFarmacologia Tarea 5 AntiparacitariosGloria Cecilia Florez GonzalezPas encore d'évaluation
- M-Epi-016 Perfil Epidemiológico de VillavicencioDocument31 pagesM-Epi-016 Perfil Epidemiológico de VillavicencioTusilueta Estetica SaludPas encore d'évaluation
- Cosmetologia: Spa ArmoníasDocument11 pagesCosmetologia: Spa ArmoníasLianexis Franco De BarretoPas encore d'évaluation
- (5.4) Semiología ObstétricaDocument4 pages(5.4) Semiología ObstétricaANA VICTORIA MARTINEZ OCHOCHOQUEPas encore d'évaluation
- Cancer de Cuello UterinoDocument4 pagesCancer de Cuello UterinoDaisy Pizarro Aquino100% (1)
- USMP Sesión 11 Salud Ocupacional Virtual 2020-IDocument32 pagesUSMP Sesión 11 Salud Ocupacional Virtual 2020-ISheyla Osorio Ramos0% (1)
- Historia ClinicaDocument5 pagesHistoria ClinicaDIANA PAOLA LUNA MENDEZPas encore d'évaluation
- Actividad 2 Módulo 2 ATENCION FARMACEUTICA 2Document2 pagesActividad 2 Módulo 2 ATENCION FARMACEUTICA 2yessica rochaPas encore d'évaluation
- METODOS DE ESTUDIO - RX.TAC - ECOCARDIOGRAFIA.ECO DOPPLER - CARDIORM-fusionadoDocument132 pagesMETODOS DE ESTUDIO - RX.TAC - ECOCARDIOGRAFIA.ECO DOPPLER - CARDIORM-fusionadoRoxana Alarcón VásquezPas encore d'évaluation
- Paquetes de Atencion Integral Del NiñoDocument20 pagesPaquetes de Atencion Integral Del NiñoWilliam Zavala Neyra67% (3)
- Valoracion 14 Necesidades v. HendersonDocument19 pagesValoracion 14 Necesidades v. HendersonViridiana IxtepanPas encore d'évaluation
- Obstetricia y GinecologiaDocument9 pagesObstetricia y GinecologiaYubetsy Melissa Tisoc HerreraPas encore d'évaluation
- Infecciones Altas y BajasDocument14 pagesInfecciones Altas y BajasLuz CacorPas encore d'évaluation
- Treatment of Primary Hypothyroidism in Adults - UpToDateDocument37 pagesTreatment of Primary Hypothyroidism in Adults - UpToDateAlison AcuranaPas encore d'évaluation
- Evaluar OteroGabilondo LuciaDocument15 pagesEvaluar OteroGabilondo LuciaCarmen SosaPas encore d'évaluation
- Semiologia DermatológicaDocument14 pagesSemiologia DermatológicaStelio ManhiquePas encore d'évaluation
- Guía 9° - 7 Función ExponencialDocument10 pagesGuía 9° - 7 Función ExponencialJulián RiveraPas encore d'évaluation
- Antropologia TrabajoDocument10 pagesAntropologia TrabajoAngel Alejandro TCPas encore d'évaluation
- RevistaDocument26 pagesRevistaMariel LivengoodPas encore d'évaluation
- Los Secretos De La Mente Millonaria: Domina el juego de la riquezaD'EverandLos Secretos De La Mente Millonaria: Domina el juego de la riquezaÉvaluation : 5 sur 5 étoiles5/5 (457)
- El Monje Que Vendio Su Ferrari: Una Fábula EspiritualD'EverandEl Monje Que Vendio Su Ferrari: Una Fábula EspiritualÉvaluation : 4.5 sur 5 étoiles4.5/5 (1700)
- Tus Zonas Erroneas: Guía Para Combatir las Causas de la InfelicidadD'EverandTus Zonas Erroneas: Guía Para Combatir las Causas de la InfelicidadÉvaluation : 4.5 sur 5 étoiles4.5/5 (1833)
- Recupera tu mente, reconquista tu vidaD'EverandRecupera tu mente, reconquista tu vidaÉvaluation : 5 sur 5 étoiles5/5 (9)
- Psicología oscura: Una guía esencial de persuasión, manipulación, engaño, control mental, negociación, conducta humana, PNL y guerra psicológicaD'EverandPsicología oscura: Una guía esencial de persuasión, manipulación, engaño, control mental, negociación, conducta humana, PNL y guerra psicológicaÉvaluation : 4.5 sur 5 étoiles4.5/5 (766)
- Yo Pude, ¡Tú Puedes!: Cómo tomar el control de tu bienestar emocional y convertirte en una persona imparable (edición revisada y expandida)D'EverandYo Pude, ¡Tú Puedes!: Cómo tomar el control de tu bienestar emocional y convertirte en una persona imparable (edición revisada y expandida)Évaluation : 5 sur 5 étoiles5/5 (9)
- Cómo hacer que te pasen cosas buenas: Entiende tu cerebro, gestiona tus emociones, mejora tu vidaD'EverandCómo hacer que te pasen cosas buenas: Entiende tu cerebro, gestiona tus emociones, mejora tu vidaÉvaluation : 5 sur 5 étoiles5/5 (1875)
- La revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaD'EverandLa revolución de la glucosa: Equilibra tus niveles de glucosa y cambiarás tu salud y tu vidaÉvaluation : 5 sur 5 étoiles5/5 (203)
- ¡Tómate un respiro! Mindfulness: El arte de mantener la calma en medio de la tempestadD'Everand¡Tómate un respiro! Mindfulness: El arte de mantener la calma en medio de la tempestadÉvaluation : 5 sur 5 étoiles5/5 (199)
- La violencia invisible: Identificar, entender y superar la violencia psicológica que sufrimos (y ejercemos) en nuestra vida cotidianaD'EverandLa violencia invisible: Identificar, entender y superar la violencia psicológica que sufrimos (y ejercemos) en nuestra vida cotidianaÉvaluation : 4 sur 5 étoiles4/5 (2)
- Resetea tu mente. Descubre de lo que eres capazD'EverandResetea tu mente. Descubre de lo que eres capazÉvaluation : 5 sur 5 étoiles5/5 (196)