Vous aimerez peut-être aussi
- Anatomia II Del CorazónDocument3 pagesAnatomia II Del CorazónJulianaleslieTJPas encore d'évaluation
- Sistema InmunologicoDocument20 pagesSistema InmunologicoJulianaleslieTJPas encore d'évaluation
- Anatomía I Del CorazónDocument3 pagesAnatomía I Del CorazónJulianaleslieTJPas encore d'évaluation
- Cardiopatía Isquémica, Crónica - Angina Estable, CoronariografíaDocument5 pagesCardiopatía Isquémica, Crónica - Angina Estable, CoronariografíaJulianaleslieTJPas encore d'évaluation
- Preguntas de Sistema InmunologicoDocument3 pagesPreguntas de Sistema InmunologicoJulianaleslieTJPas encore d'évaluation
- Tejido Nervioso2Document19 pagesTejido Nervioso2JulianaleslieTJPas encore d'évaluation
- Hiperplasia Prostatica BenignaDocument19 pagesHiperplasia Prostatica BenignaJulianaleslieTJPas encore d'évaluation
- Fisologi de La Miccion y Urodinamia y Patologias VesicalesDocument57 pagesFisologi de La Miccion y Urodinamia y Patologias VesicalesJulianaleslieTJPas encore d'évaluation
- Aparato UrinarioDocument6 pagesAparato UrinarioJulianaleslieTJPas encore d'évaluation
- Incontinencia Urinaria de EsfuerzoDocument34 pagesIncontinencia Urinaria de EsfuerzoJulianaleslieTJPas encore d'évaluation
- BIOLOGIApreDocument108 pagesBIOLOGIApreJulianaleslieTJPas encore d'évaluation
- ORGANOGRAFÍA VEGETAL II 1roDocument5 pagesORGANOGRAFÍA VEGETAL II 1roJulianaleslieTJPas encore d'évaluation
- Preguntas de Biologia CelularDocument3 pagesPreguntas de Biologia CelularJulianaleslieTJPas encore d'évaluation
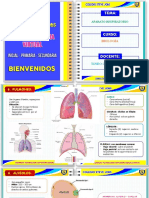
- APARATO RESPIRATORIO - PulmonesDocument4 pagesAPARATO RESPIRATORIO - PulmonesJulianaleslieTJPas encore d'évaluation
- Aplicacón de Calculo Iii A La Ingenieria AmbientalDocument4 pagesAplicacón de Calculo Iii A La Ingenieria AmbientalJulianaleslieTJPas encore d'évaluation
- Examenes de La UnhevalDocument10 pagesExamenes de La UnhevalJulianaleslieTJPas encore d'évaluation
- GERENCIADocument4 pagesGERENCIAJulianaleslieTJPas encore d'évaluation
- Principios Activos Aloe VeraDocument21 pagesPrincipios Activos Aloe VeraJoselitoPas encore d'évaluation
- Informe #9 - Química Orgánica II PDFDocument26 pagesInforme #9 - Química Orgánica II PDFDevra GomezPas encore d'évaluation
- 5 CLASE SEMANA 5-Importancia de Los MacronutrientesDocument60 pages5 CLASE SEMANA 5-Importancia de Los MacronutrientesJYOSEF GAVILAN HUAMANPas encore d'évaluation
- Universidad Iberoamericana (UNIBE)Document7 pagesUniversidad Iberoamericana (UNIBE)Mariadelcarmen Garabito Sierra0% (1)
- Wuolah Free PREPARACION DEL EXAMEN TEMA 3Document7 pagesWuolah Free PREPARACION DEL EXAMEN TEMA 3Ithaisa Agustina Rodríguez PérezPas encore d'évaluation
- Las Proteínas: Docente: IntegrantesDocument66 pagesLas Proteínas: Docente: Integrantespatricia lourdes PimentelPas encore d'évaluation
- Experimento Del Sistema DigestivoDocument12 pagesExperimento Del Sistema DigestivoElizabeth CrisantoPas encore d'évaluation
- Planta Moringa Aminoacidos EscencialesDocument8 pagesPlanta Moringa Aminoacidos EscencialesLuis Miguel Marquez YepesPas encore d'évaluation
- TALLER Repaso AA PEPTIDOS ProteinasDocument2 pagesTALLER Repaso AA PEPTIDOS ProteinasTatiana ArteagaPas encore d'évaluation
- AminoácidosDocument16 pagesAminoácidosEmilyPas encore d'évaluation
- Informe Tectnico Isabion PDFDocument52 pagesInforme Tectnico Isabion PDFJavier Cobos MoraPas encore d'évaluation
- Curvas de Titulación de AminoácidosDocument2 pagesCurvas de Titulación de AminoácidosFernanda DomínguezPas encore d'évaluation
- Métodos Evaluación de La ProteínaDocument1 pageMétodos Evaluación de La ProteínaIlanit MeierPas encore d'évaluation
- Folleto 3 - BCDocument19 pagesFolleto 3 - BCAnonymous PG6kaDl22EPas encore d'évaluation
- AdnDocument3 pagesAdnAdriana AbadPas encore d'évaluation
- Anotación de ProteínasDocument8 pagesAnotación de Proteínasfelipe romeroPas encore d'évaluation
- Asaduroglu Nutricion y Alimentacion HumanaDocument145 pagesAsaduroglu Nutricion y Alimentacion HumanaSofiaPas encore d'évaluation
- BioquimicaDocument126 pagesBioquimicaSousuke SagaraPas encore d'évaluation
- Proteinas Quimica ExpoDocument25 pagesProteinas Quimica ExpoMaría Claudia Cáceres ObadaPas encore d'évaluation
- BioquimicaDocument11 pagesBioquimicaTaatiiaanaa CastillooPas encore d'évaluation
- Practica 4 BiocaDocument12 pagesPractica 4 BiocaVáleri Alexandra100% (1)
- AlcachofaDocument17 pagesAlcachofaJulissa LibertadPas encore d'évaluation
- %plumasDocument13 pages%plumasHéctor Medina OviedoPas encore d'évaluation
- Glosario BioquimicoDocument12 pagesGlosario BioquimicoAngelitho Muñoz ChavezPas encore d'évaluation
- Taller Bioquímica 1Document17 pagesTaller Bioquímica 1manuelaPas encore d'évaluation
- Unidadii Aminoacidos Peptidos Proteinas 21268 PDFDocument52 pagesUnidadii Aminoacidos Peptidos Proteinas 21268 PDFColín Poblete BaezaPas encore d'évaluation
- Repartido 1 Aminoácidos y ProteínasDocument1 pageRepartido 1 Aminoácidos y ProteínasgPas encore d'évaluation
- Estructura y Composición Quimica de La PielDocument13 pagesEstructura y Composición Quimica de La PielYampier LopezPas encore d'évaluation
- Qué Es Un AminoácidoDocument4 pagesQué Es Un AminoácidoYiye PadillaPas encore d'évaluation
- Determinación de Proteínas Lautaro SchisslerDocument9 pagesDeterminación de Proteínas Lautaro SchisslerLautaro SchisslerPas encore d'évaluation