Vous aimerez peut-être aussi
- La Membrana CelularDocument68 pagesLa Membrana CelularMaría Ángel DoriaPas encore d'évaluation
- Estructura y función de la membrana plasmáticaDocument30 pagesEstructura y función de la membrana plasmáticaRadar100% (2)
- Transporte de ProteinasDocument9 pagesTransporte de ProteinasGise MathierPas encore d'évaluation
- 8 Ribosomas y OrganelosDocument47 pages8 Ribosomas y OrganelosShandu RkPas encore d'évaluation
- Membrana CitoplasmaticaDocument47 pagesMembrana CitoplasmaticaFernando MartinezPas encore d'évaluation
- La Membrana CelularDocument32 pagesLa Membrana CelularMaycol Huaman CastilloPas encore d'évaluation
- Membrana celular: Transporte a través de la membranaDocument4 pagesMembrana celular: Transporte a través de la membranaDiego AgudeloPas encore d'évaluation
- Semana 07 Terminado Expo BioDocument4 pagesSemana 07 Terminado Expo BioSofiaPas encore d'évaluation
- Transporte celular y membrana plasmáticaDocument4 pagesTransporte celular y membrana plasmáticaSandra Valentina100% (1)
- Funciones Del AmpcDocument3 pagesFunciones Del AmpcRaulPas encore d'évaluation
- Membrana Celular.Document15 pagesMembrana Celular.Paul Berrones100% (1)
- Integrinas y selectinas: moléculas de adhesión celularDocument2 pagesIntegrinas y selectinas: moléculas de adhesión celularJose Eduardo LopezPas encore d'évaluation
- De Gen A Proteina-Inf11-SemDocument7 pagesDe Gen A Proteina-Inf11-SemLuis Alejandro Quispe PoncePas encore d'évaluation
- Citoesqueleto: FuncionesDocument14 pagesCitoesqueleto: Funcioneskarina ramosPas encore d'évaluation
- Metabolismo y Respiración CelularDocument26 pagesMetabolismo y Respiración CelularHerbert JaimesPas encore d'évaluation
- Tejido ConectivoDocument27 pagesTejido ConectivoAle univallePas encore d'évaluation
- Recepción Celular. Mecanismos MolecularesDocument17 pagesRecepción Celular. Mecanismos MolecularesTEFAPas encore d'évaluation
- Moléculas de relación celular: integrinas, cadherinas y selectinasDocument6 pagesMoléculas de relación celular: integrinas, cadherinas y selectinasXiomara EvangelistaPas encore d'évaluation
- BioelectricidadDocument32 pagesBioelectricidadMaritho Vera SànchezPas encore d'évaluation
- Histologia Arterias Venas y CapilaresDocument45 pagesHistologia Arterias Venas y CapilaresJorge Bautistad100% (3)
- Señalización Celular - Matriz ExtracelularDocument7 pagesSeñalización Celular - Matriz ExtracelularEnzo Guillermo RamirezPas encore d'évaluation
- Actividad EnzimáticaDocument11 pagesActividad EnzimáticaEnmanuel Samudio100% (1)
- MICROFILAMENTOSDocument7 pagesMICROFILAMENTOSalmanza_1973Pas encore d'évaluation
- Placenta MacroscópicaDocument11 pagesPlacenta MacroscópicaLiliana Julissa García AlarcónPas encore d'évaluation
- Membrana Plasmática CUESTIONARIODocument9 pagesMembrana Plasmática CUESTIONARIOClifor Ivan Cuello100% (1)
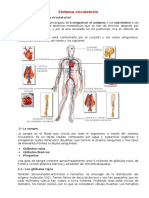
- Sistema CirculatorioDocument6 pagesSistema CirculatorioWalter ParedesPas encore d'évaluation
- Inmunologia en El EmbarazoDocument36 pagesInmunologia en El EmbarazoDioser AvilaPas encore d'évaluation
- Estructura y Función de La Membrana PlasmáticaDocument102 pagesEstructura y Función de La Membrana Plasmátical@sm@rinos1100% (1)
- Citoesqueleto, ResumenDocument13 pagesCitoesqueleto, ResumenMKsander GarcíaPas encore d'évaluation
- Composición y Organización Molecular de La Membrana CelularDocument13 pagesComposición y Organización Molecular de La Membrana CelularAlexandra Torres Basurto0% (1)
- Senalizacion CelularDocument20 pagesSenalizacion CelularWilmer HuamaniPas encore d'évaluation
- Membrana PlasmaticaDocument53 pagesMembrana PlasmaticavictoriaPas encore d'évaluation
- Reproduccion y Muerte CelularDocument18 pagesReproduccion y Muerte CelularRobert Carvajal0% (1)
- Informe 12 - Informatica IDocument9 pagesInforme 12 - Informatica IHector Arbañil NanfuñayPas encore d'évaluation
- Biología-Sem-InformeDocument10 pagesBiología-Sem-InformeXiomara Janne Linares SaldañaPas encore d'évaluation
- La Membrana CelularDocument22 pagesLa Membrana CelularJorge AndrésPas encore d'évaluation
- Enzimas. Conceptos BasicosDocument3 pagesEnzimas. Conceptos BasicosJess QuijadaPas encore d'évaluation
- Endosomas y LisosomasDocument24 pagesEndosomas y LisosomasAurelia Gonzales Calle93% (14)
- Segundos MensajerosDocument14 pagesSegundos MensajerosArnold Gomez AlvaPas encore d'évaluation
- Observación de embriones: AbdomenDocument7 pagesObservación de embriones: AbdomenGustavo Herrera MontenegroPas encore d'évaluation
- 8° Guía 3 Organelos CelularesDocument3 pages8° Guía 3 Organelos Celularescesar villaPas encore d'évaluation
- Glosario Biología MolecularDocument7 pagesGlosario Biología MolecularValeria100% (1)
- Uniones Celulares Seminario 11Document29 pagesUniones Celulares Seminario 11xiomara EusebioPas encore d'évaluation
- Glosario Membrana CelularDocument3 pagesGlosario Membrana CelularNicole OrjuelaPas encore d'évaluation
- ComCelMensQuímDocument25 pagesComCelMensQuímVictoria Sepulveda Salazar100% (1)
- Tejido EpitelialDocument6 pagesTejido Epitelialktrance_Pas encore d'évaluation
- Especialización de La MembranaDocument2 pagesEspecialización de La MembranaMarisol100% (2)
- Transporte A Través de La Membrana CelularesDocument8 pagesTransporte A Través de La Membrana CelularesNando MéridaPas encore d'évaluation
- Farmacología de la acetilcolina en el sistema nerviosoDocument14 pagesFarmacología de la acetilcolina en el sistema nerviosoHija de DiosPas encore d'évaluation
- Transporte de proteínas en la célulaDocument4 pagesTransporte de proteínas en la célulaEsteban RuedaPas encore d'évaluation
- Respiración Celular (Guia)Document33 pagesRespiración Celular (Guia)Lourdes PerdomoPas encore d'évaluation
- Clase 18 Membrana CelularDocument9 pagesClase 18 Membrana Celularduartec7Pas encore d'évaluation
- Transporte ActivoDocument16 pagesTransporte ActivoSofia EspinozaPas encore d'évaluation
- Informe Práctica S10Document10 pagesInforme Práctica S10Alexander E. Bruckmann AvilaPas encore d'évaluation
- Capitul Iii y IvDocument33 pagesCapitul Iii y IvAracely PilamontaPas encore d'évaluation
- 3.permeabilidad de La MembranaDocument20 pages3.permeabilidad de La MembranaJunior Rey ColomePas encore d'évaluation
- Membrana CelularDocument26 pagesMembrana CelularJulio Ricardo Ruiz CardonaPas encore d'évaluation
- Capitulo 7 Estructura y Funcion de La MembranaDocument4 pagesCapitulo 7 Estructura y Funcion de La MembranaRicardo PosadasPas encore d'évaluation
- Membrana CelularDocument4 pagesMembrana CelularMichelle Estefanía Arredondo ArmasPas encore d'évaluation
- Resumen Membrana PlasmáticaDocument4 pagesResumen Membrana PlasmáticaALESSANDRO JOSUE ALVARADO GALEANOPas encore d'évaluation
- Celula PDFDocument24 pagesCelula PDFAndrea ChinchillaPas encore d'évaluation
- La Excitabilidad Neuronal y Los Canales de PotasioDocument7 pagesLa Excitabilidad Neuronal y Los Canales de PotasioJohn Eduardo Henriquez BorbonPas encore d'évaluation
- La MielinaDocument5 pagesLa MielinaF1101Pas encore d'évaluation
- Manual DemenciaDocument187 pagesManual DemenciaF1101Pas encore d'évaluation
- Insectos y Aracnidos Beneficiosos-Para AgriculturaDocument6 pagesInsectos y Aracnidos Beneficiosos-Para AgriculturaF1101Pas encore d'évaluation
- Fertilizacion en Citricultura EcologicaDocument26 pagesFertilizacion en Citricultura EcologicaOswaldo Suri QuispePas encore d'évaluation
- Seminar I o SueloDocument26 pagesSeminar I o Sueloricardot8aPas encore d'évaluation
- La Excitabilidad Neuronal y Los Canales de PotasioDocument7 pagesLa Excitabilidad Neuronal y Los Canales de PotasioJohn Eduardo Henriquez BorbonPas encore d'évaluation
- Fisiología del SN: Impulso nervioso y sinapsisDocument9 pagesFisiología del SN: Impulso nervioso y sinapsisF1101Pas encore d'évaluation
- Manual DemenciaDocument187 pagesManual DemenciaF1101Pas encore d'évaluation
- Fauna SilvestreDocument123 pagesFauna SilvestreF1101Pas encore d'évaluation
- Mundo Mapa 04 PDFDocument1 pageMundo Mapa 04 PDFatila117Pas encore d'évaluation
- Manual DemenciaDocument187 pagesManual DemenciaF1101Pas encore d'évaluation
- Tema 5Document10 pagesTema 5Gaby BuPas encore d'évaluation
- Tema 5Document10 pagesTema 5Gaby BuPas encore d'évaluation
- Narracion EjemplosDocument6 pagesNarracion EjemplosF1101Pas encore d'évaluation
- MatematicosDocument4 pagesMatematicosLeydi Esther Ventura RojasPas encore d'évaluation
- Matematica FinancieraDocument24 pagesMatematica FinancieraF1101Pas encore d'évaluation
- Texto NarrativoDocument7 pagesTexto Narrativojosh bernalPas encore d'évaluation
- Análisis de datos estadísticos descriptivosDocument10 pagesAnálisis de datos estadísticos descriptivosrobbertfrippPas encore d'évaluation
- 6 - Proceso Quirúrgico 2015 V - 2Document46 pages6 - Proceso Quirúrgico 2015 V - 2Daner Vera FuenzalidaPas encore d'évaluation
- MatematicosDocument4 pagesMatematicosLeydi Esther Ventura RojasPas encore d'évaluation
- Ejercicios (Con Solución) de EstadísticaDocument11 pagesEjercicios (Con Solución) de EstadísticaISABEL69% (13)
- Ejercicios (Con Solución) de EstadísticaDocument11 pagesEjercicios (Con Solución) de EstadísticaISABEL69% (13)
- Actividades Recuperacion 4o ESOBiologia2017Document18 pagesActividades Recuperacion 4o ESOBiologia2017maria100% (1)
- Marcadores MitocondrialesDocument10 pagesMarcadores MitocondrialesF1101Pas encore d'évaluation
- Examen Corregido BiologiaDocument12 pagesExamen Corregido BiologiaF1101Pas encore d'évaluation
- Marcadores MitocondrialesDocument10 pagesMarcadores MitocondrialesF1101Pas encore d'évaluation
- Genoma MitocondrialDocument83 pagesGenoma MitocondrialF1101Pas encore d'évaluation
- Aplicaciones Geometria AnaliticaDocument3 pagesAplicaciones Geometria AnaliticaSandra Gabriela Coca OrtizPas encore d'évaluation
- TFG Rafael De-Solis Galindo 2018 PDFDocument101 pagesTFG Rafael De-Solis Galindo 2018 PDFDaniielaPérezPas encore d'évaluation
- Diseño Malla PAT 2013Document12 pagesDiseño Malla PAT 2013Raul LopezPas encore d'évaluation
- Informe 1 Metodo DumasDocument8 pagesInforme 1 Metodo DumasAnita UshiñaPas encore d'évaluation
- KMKPDocument17 pagesKMKPMiguel Murga MirandaPas encore d'évaluation
- 6 Programa de Uso Eficiente y Ahorro de Energia PUEAE PDFDocument26 pages6 Programa de Uso Eficiente y Ahorro de Energia PUEAE PDFPedro Antonio PaniaguaPas encore d'évaluation
- LEVANTAMIENTO CATASTRAL Oscar Arotaipe GutierrezDocument46 pagesLEVANTAMIENTO CATASTRAL Oscar Arotaipe GutierrezOscar MartinezPas encore d'évaluation
- 02 - Conversion de UnidadesDocument6 pages02 - Conversion de UnidadesGonzalo GonzaloPas encore d'évaluation
- La Estructura Del Fullereno C60 y Sus AplicacionesDocument3 pagesLa Estructura Del Fullereno C60 y Sus AplicacionesAraceliPas encore d'évaluation
- Costos y ventajas del sistema Raise Borer para perforación verticalDocument9 pagesCostos y ventajas del sistema Raise Borer para perforación verticalAndresCernaAPas encore d'évaluation
- Guía de Laboratoriode Orgámica IaDocument29 pagesGuía de Laboratoriode Orgámica IaLeonardo Valderrama PenaPas encore d'évaluation
- Ejercicios Resueltos Ley de KirchhoffDocument7 pagesEjercicios Resueltos Ley de KirchhoffJean Carlos Mesa Guarin100% (4)
- Ecuaciones Diferenciales en La Ingeniería en ComputaciónDocument6 pagesEcuaciones Diferenciales en La Ingeniería en ComputaciónUlises Landázuri Brambila0% (1)
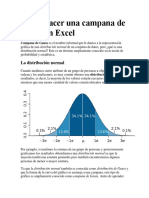
- Cómo Hacer Una Campana de Gauss en ExcelDocument5 pagesCómo Hacer Una Campana de Gauss en ExcelROY EDSON ARPASI BARTOLOPas encore d'évaluation
- Fisica Ii - Lab 1 PDFDocument15 pagesFisica Ii - Lab 1 PDFrianiPas encore d'évaluation
- Aplicaciones de las matrices en modelos de crecimiento poblacionalDocument9 pagesAplicaciones de las matrices en modelos de crecimiento poblacionaldegelbbis_29_1@hotmail.com0% (1)
- Act 5 - QuizDocument7 pagesAct 5 - QuizJean Paul RossiPas encore d'évaluation
- Practica N°02 - Mecanica de Fluidos - HidrostaticaDocument12 pagesPractica N°02 - Mecanica de Fluidos - HidrostaticaOscar Adrian Chavez PanduroPas encore d'évaluation
- TransformadoresDocument46 pagesTransformadoresAbraham Galindo FloresPas encore d'évaluation
- Choques ExperimentosDocument7 pagesChoques ExperimentosFaber KingPas encore d'évaluation
- Apuntes. ProbabilidadDocument2 pagesApuntes. ProbabilidadMaria JosePas encore d'évaluation
- UV VisDocument9 pagesUV VisJonatan DavidPas encore d'évaluation
- Conferencia Construccion de Tuneles en Suelos y Rocas Por AnDocument110 pagesConferencia Construccion de Tuneles en Suelos y Rocas Por AnIng Ignacio Rojo Gastelum100% (1)
- 7 EnergíaDocument16 pages7 Energíafrank xd0% (1)
- Intervalos de datosDocument19 pagesIntervalos de datosDaisy Alejandra Muñoz SilvaPas encore d'évaluation
- POTHENOTDocument14 pagesPOTHENOTVictor Nina50% (2)
- Biografias de MatematicosDocument79 pagesBiografias de MatematicosLuis AngelPas encore d'évaluation
- Taller Completacion de PozosDocument13 pagesTaller Completacion de PozosVariedadesLuPas encore d'évaluation
- Electonica de PotenciaDocument6 pagesElectonica de PotenciaVictor Cerda MorenoPas encore d'évaluation
- Ejercicios Física 3 WMDocument9 pagesEjercicios Física 3 WMvivianPas encore d'évaluation