Vous aimerez peut-être aussi
- Academic Job Interviews: Questions and AdviceDocument26 pagesAcademic Job Interviews: Questions and AdviceAstarag DasPas encore d'évaluation
- Conn - ResumeDocument7 pagesConn - ResumeAstarag DasPas encore d'évaluation
- 15 Toughest Interview Questions and Answers!: 1. Why Do You Want To Work in This Industry?Document8 pages15 Toughest Interview Questions and Answers!: 1. Why Do You Want To Work in This Industry?johnlemPas encore d'évaluation
- PselectinDocument7 pagesPselectinAstarag DasPas encore d'évaluation
- Office 2003 Updates-WebsitesDocument1 pageOffice 2003 Updates-WebsitesAstarag DasPas encore d'évaluation
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
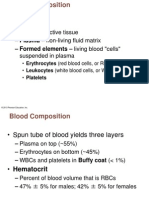
- Physiology, Lecture 5, Blood (Lecture Notes)Document10 pagesPhysiology, Lecture 5, Blood (Lecture Notes)Ali Al-Qudsi100% (1)
- 03f - 8-06 Awake ImagesDocument2 pages03f - 8-06 Awake ImagesRobert CamachoPas encore d'évaluation
- Blood RecreationDocument387 pagesBlood RecreationAlexandr TrotskyPas encore d'évaluation
- Assessment of Inflammatory Markers in Otosclerosis PatientsDocument5 pagesAssessment of Inflammatory Markers in Otosclerosis PatientsCinthia Marissa Arellano GarcíaPas encore d'évaluation
- Case Study - Dengue FeverDocument61 pagesCase Study - Dengue FeverFrancis AdrianPas encore d'évaluation
- CardioVascular and Interventional RadiologyDocument233 pagesCardioVascular and Interventional RadiologyCristi Arias BadaPas encore d'évaluation
- Patient Evaluat-WPS OfficeDocument109 pagesPatient Evaluat-WPS OfficeCounselor Optimus EduconPas encore d'évaluation
- Unit 5 Blood TransfusionDocument10 pagesUnit 5 Blood TransfusionMarianne Gonzales-HerreraPas encore d'évaluation
- 4.1 Blood Component SeparationDocument8 pages4.1 Blood Component Separationayan CustodyPas encore d'évaluation
- Quantitative Platelet Disorder: Hannie M. SacatanDocument24 pagesQuantitative Platelet Disorder: Hannie M. SacatanFearless AngelPas encore d'évaluation
- Principles of Wound HealingDocument28 pagesPrinciples of Wound HealingRafael BagusPas encore d'évaluation
- 4 Safe Transfusion Right Blood Right Patient Right Time and Right PlaceDocument11 pages4 Safe Transfusion Right Blood Right Patient Right Time and Right PlaceEtrya WiandaPas encore d'évaluation
- Fenwal-Product Catalog v2010Document36 pagesFenwal-Product Catalog v2010Campaign Media0% (1)
- Hematology - Steininger ReviewDocument30 pagesHematology - Steininger ReviewIssa AlejoPas encore d'évaluation
- HipersplenismDocument6 pagesHipersplenismDany YayanPas encore d'évaluation
- Plasma Separation Review PPT 2Document21 pagesPlasma Separation Review PPT 2NhhjPas encore d'évaluation
- Yoon 2014Document7 pagesYoon 2014aleegaleanoPas encore d'évaluation
- BCC3000 Service ManualDocument54 pagesBCC3000 Service Manualzelika Jemal78% (9)
- Blood Lecture PowerpointDocument116 pagesBlood Lecture Powerpointdrmarybeth100% (3)
- Anatomy Final Exam Study GuideDocument15 pagesAnatomy Final Exam Study GuideAndy Tran100% (3)
- Blood Chem QuizDocument3 pagesBlood Chem QuizJaycarlo SaltivanPas encore d'évaluation
- Hemophilia Powerpoint-Kuliah FKIK UNTANDocument33 pagesHemophilia Powerpoint-Kuliah FKIK UNTANAgung TriatmojoPas encore d'évaluation
- Anticoagulants DaniellewelschmeyerDocument30 pagesAnticoagulants Daniellewelschmeyersumanth gowdaPas encore d'évaluation
- Hemostasis in Dentistry PDFDocument204 pagesHemostasis in Dentistry PDFMeko GegechkoriPas encore d'évaluation
- Complications To PhlebotomyDocument4 pagesComplications To Phlebotomybenrmt67% (3)
- Hematologic System, Oncologic Disorders & Anemias: DR Ibrahreem Bashayreh, RN, PHDDocument68 pagesHematologic System, Oncologic Disorders & Anemias: DR Ibrahreem Bashayreh, RN, PHDYA HOPas encore d'évaluation
- Immune Thrombocytopenia (ITP) : Signs and SymptomsDocument35 pagesImmune Thrombocytopenia (ITP) : Signs and SymptomsRachmad SammuliaPas encore d'évaluation
- TropocellsDocument6 pagesTropocellsIsrael Exporter100% (1)
- Thrombotic Thrombocytopenic Purpura, Heparin-Induced Thrombocytopenia, and Disseminated Intravascular Coagulation 2020Document21 pagesThrombotic Thrombocytopenic Purpura, Heparin-Induced Thrombocytopenia, and Disseminated Intravascular Coagulation 2020Cris TobalPas encore d'évaluation
- Hema 2 - Prelim Topic 3 - Platelet Disorders and Laboratory TestsDocument8 pagesHema 2 - Prelim Topic 3 - Platelet Disorders and Laboratory TestsLowenstein JenzenPas encore d'évaluation