Vous aimerez peut-être aussi
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (120)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (399)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- 6th Central Pay Commission Salary CalculatorDocument15 pages6th Central Pay Commission Salary Calculatorrakhonde100% (436)
- Basic of AccupressureDocument48 pagesBasic of AccupressureRavikumarrd Ravi100% (2)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- Cranial Nerves NeuroanatomyDocument57 pagesCranial Nerves NeuroanatomyArvin TanPas encore d'évaluation
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- Vein Vs ArteryDocument4 pagesVein Vs ArterySaarwin MuruganPas encore d'évaluation
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (73)
- Open Bites and Tongue PositionDocument26 pagesOpen Bites and Tongue PositionDimitris GalaktopoulosPas encore d'évaluation
- Functional Organization of Cardiovascular SystemDocument19 pagesFunctional Organization of Cardiovascular SystemIbtesam Mohammed100% (2)
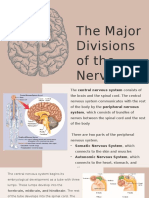
- The Major Divisions of The Nervous SystemDocument17 pagesThe Major Divisions of The Nervous SystemLiana BaluyotPas encore d'évaluation
- Nirc PDFDocument160 pagesNirc PDFCharlie Magne G. Santiaguel100% (3)
- Zoology MCQDocument15 pagesZoology MCQeesha iqbal67% (3)
- Rice Husk Gas Stove Handbook - Alexis T.BelonioDocument155 pagesRice Husk Gas Stove Handbook - Alexis T.BelonioAris TrihandokoPas encore d'évaluation
- Prinsip Primary SurveyDocument41 pagesPrinsip Primary SurveyTRI WAHYUNINGSIHPas encore d'évaluation
- Periodontitis Symptoms Treatment and Prevention Rosemarie E. WalchuckDocument286 pagesPeriodontitis Symptoms Treatment and Prevention Rosemarie E. WalchuckDaniel RapcauPas encore d'évaluation
- Laporan Kasus PPOKDocument23 pagesLaporan Kasus PPOKbahrinaPas encore d'évaluation
- Commissioner of Internal Renenue V San Roque Power CorporationDocument49 pagesCommissioner of Internal Renenue V San Roque Power Corporationtristram04Pas encore d'évaluation
- Ra 9700Document14 pagesRa 9700tristram04Pas encore d'évaluation
- Provincial Assessor of Cagayan, CBAA Case No. L-03Document5 pagesProvincial Assessor of Cagayan, CBAA Case No. L-03tristram04Pas encore d'évaluation
- Doing Business in The PhilippinesDocument0 pageDoing Business in The Philippinestristram04Pas encore d'évaluation
- Egal and Udicial Thics: Yllabi For AR XaminationsDocument4 pagesEgal and Udicial Thics: Yllabi For AR Xaminationscmv mendozaPas encore d'évaluation
- ION 17Q Sep2013Document37 pagesION 17Q Sep2013tristram04Pas encore d'évaluation
- Dela Paz V CADocument6 pagesDela Paz V CAtristram04Pas encore d'évaluation
- AAA 17Q Jun2010Document27 pagesAAA 17Q Jun2010tristram04Pas encore d'évaluation
- Austrian Economics Booms in Popularity Busts Mainstream MythsDocument4 pagesAustrian Economics Booms in Popularity Busts Mainstream Mythstristram04Pas encore d'évaluation
- Drug Study Vent Forgram)Document2 pagesDrug Study Vent Forgram)arvie_mirandaPas encore d'évaluation
- Kangkong (Ipomoea Aquatica) : A Wriggler Killer: An Investigatory Project Presented ToDocument11 pagesKangkong (Ipomoea Aquatica) : A Wriggler Killer: An Investigatory Project Presented ToAries Palao Austria100% (1)
- Module 9 Part A: Monitoring and Evaluation: Image Courtesy Of: World Lung FoundationDocument41 pagesModule 9 Part A: Monitoring and Evaluation: Image Courtesy Of: World Lung FoundationMataqaana? GroupPas encore d'évaluation
- Topographic Anatomy of Abdominal OrgansDocument18 pagesTopographic Anatomy of Abdominal OrgansHasnain IdreesPas encore d'évaluation
- Submental Artery Island Flap Technique For Head Neck ReconstructionDocument18 pagesSubmental Artery Island Flap Technique For Head Neck ReconstructionMahir NuredinPas encore d'évaluation
- Actividad y Traducciones de Terminos Medicos + AdjetivosDocument2 pagesActividad y Traducciones de Terminos Medicos + AdjetivosJosue CastilloPas encore d'évaluation
- Fluid and Electrolytes, Balance and DisturbancesDocument157 pagesFluid and Electrolytes, Balance and DisturbancesDani PhilipPas encore d'évaluation
- 03etiology and Pathogenesis of Neonatal EncephalopathyDocument22 pages03etiology and Pathogenesis of Neonatal EncephalopathyDaniela MuñozPas encore d'évaluation
- Halosil Athletics Sell Sheet 060817 PDFDocument2 pagesHalosil Athletics Sell Sheet 060817 PDFduna tarimaPas encore d'évaluation
- Cialis Tablets in Quetta - Tadalafil Cialis - Cialis 20mgDocument4 pagesCialis Tablets in Quetta - Tadalafil Cialis - Cialis 20mgSana KhanPas encore d'évaluation
- Biostat Post TestDocument5 pagesBiostat Post TestCstive Valenzuela100% (2)
- Chemical and Nervous ControlDocument5 pagesChemical and Nervous ControlNico BereberPas encore d'évaluation
- One Hundred Magic-UsersDocument4 pagesOne Hundred Magic-UsershigherdepthsPas encore d'évaluation
- NPI and Health TeachingDocument4 pagesNPI and Health TeachingJanelle Cabida SupnadPas encore d'évaluation
- Gynae 64Document14 pagesGynae 64Jha K SujitPas encore d'évaluation
- Discurssive Writing TopicDocument8 pagesDiscurssive Writing TopicAscensión Villalba VaronaPas encore d'évaluation
- 4 Hodges and Tucker. Moving Differently in Pain. 2011Document9 pages4 Hodges and Tucker. Moving Differently in Pain. 2011ningaxshopPas encore d'évaluation
- Prosthodontics IIIDocument8 pagesProsthodontics IIICyril Almario CunananPas encore d'évaluation
- Vaginal Examination Wit RationaleDocument1 pageVaginal Examination Wit RationaleAna Maria Teresa H RollanPas encore d'évaluation
- Weekly Bedah Pediatri Minggu Ke 2 EditedDocument83 pagesWeekly Bedah Pediatri Minggu Ke 2 EditedPraditya Septian AbdulganiPas encore d'évaluation