Vous aimerez peut-être aussi
- Blood Brain Barrier Failure As A Core Mechanism in Cerebral - 2017 - AlzheimerDocument10 pagesBlood Brain Barrier Failure As A Core Mechanism in Cerebral - 2017 - AlzheimerLuis Gerardo Escandon AlcazarPas encore d'évaluation
- PhDthesisMVPhyperrefs PDFDocument191 pagesPhDthesisMVPhyperrefs PDFLuis Gerardo Escandon AlcazarPas encore d'évaluation
- Schumann Resonance and Brain Waves: A Quantum Description: Neuroquantology May 2015Document10 pagesSchumann Resonance and Brain Waves: A Quantum Description: Neuroquantology May 2015Luis Gerardo Escandon AlcazarPas encore d'évaluation
- Notes On Integration and Measure: P. B. Kronheimer, For Math 114Document27 pagesNotes On Integration and Measure: P. B. Kronheimer, For Math 114Luis Gerardo Escandon AlcazarPas encore d'évaluation
- Lec10Document5 pagesLec10Luis Gerardo Escandon AlcazarPas encore d'évaluation
- Effect of A 125 MT Static Magnetic Field On The Kinetics of Voltage Activated Na Channels in GH3 CellsDocument7 pagesEffect of A 125 MT Static Magnetic Field On The Kinetics of Voltage Activated Na Channels in GH3 CellsLuis Gerardo Escandon AlcazarPas encore d'évaluation
- Child Sex Abusers Elude Flimsy Digital Safeguards: From Dissent To Rallying Cry in State DeptDocument7 pagesChild Sex Abusers Elude Flimsy Digital Safeguards: From Dissent To Rallying Cry in State DeptLuis Gerardo Escandon AlcazarPas encore d'évaluation
- Review Essay: Transactional Man: Teaching Negotiation Strategy in The Age of TrumpDocument15 pagesReview Essay: Transactional Man: Teaching Negotiation Strategy in The Age of TrumpLuis Gerardo Escandon AlcazarPas encore d'évaluation
- Electroencephalography and Clinical N Europhysloloy (VDocument2 pagesElectroencephalography and Clinical N Europhysloloy (VLuis Gerardo Escandon AlcazarPas encore d'évaluation
- BiomagnetismDocument13 pagesBiomagnetismLuis Gerardo Escandon AlcazarPas encore d'évaluation
- Syllabus 2325 Sebahar Spring 2013Document6 pagesSyllabus 2325 Sebahar Spring 2013Luis Gerardo Escandon AlcazarPas encore d'évaluation
- Crocodile ChemistryDocument282 pagesCrocodile ChemistryLuis Gerardo Escandon AlcazarPas encore d'évaluation
- String Theory DemystifiedDocument10 pagesString Theory DemystifiedLuis Gerardo Escandon AlcazarPas encore d'évaluation
- The Einstein-Myth and The Crisis in Modern PhysicsDocument17 pagesThe Einstein-Myth and The Crisis in Modern PhysicsLuis Gerardo Escandon AlcazarPas encore d'évaluation
- Glossary of Physical Oceanography and Related Disciplines - Steven K. BaumDocument517 pagesGlossary of Physical Oceanography and Related Disciplines - Steven K. BaumLuis Gerardo Escandon Alcazar100% (1)
- What Lies Beneath: by Dan FalkDocument6 pagesWhat Lies Beneath: by Dan FalkLuis Gerardo Escandon AlcazarPas encore d'évaluation
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceD'EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceÉvaluation : 4 sur 5 étoiles4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeD'EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeÉvaluation : 4 sur 5 étoiles4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeD'EverandShoe Dog: A Memoir by the Creator of NikeÉvaluation : 4.5 sur 5 étoiles4.5/5 (537)
- Grit: The Power of Passion and PerseveranceD'EverandGrit: The Power of Passion and PerseveranceÉvaluation : 4 sur 5 étoiles4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)D'EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Évaluation : 4 sur 5 étoiles4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingD'EverandThe Little Book of Hygge: Danish Secrets to Happy LivingÉvaluation : 3.5 sur 5 étoiles3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItD'EverandNever Split the Difference: Negotiating As If Your Life Depended On ItÉvaluation : 4.5 sur 5 étoiles4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureD'EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureÉvaluation : 4.5 sur 5 étoiles4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryD'EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryÉvaluation : 3.5 sur 5 étoiles3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerD'EverandThe Emperor of All Maladies: A Biography of CancerÉvaluation : 4.5 sur 5 étoiles4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaD'EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaÉvaluation : 4.5 sur 5 étoiles4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersD'EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersÉvaluation : 4.5 sur 5 étoiles4.5/5 (345)
- On Fire: The (Burning) Case for a Green New DealD'EverandOn Fire: The (Burning) Case for a Green New DealÉvaluation : 4 sur 5 étoiles4/5 (74)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyD'EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyÉvaluation : 3.5 sur 5 étoiles3.5/5 (2259)
- Team of Rivals: The Political Genius of Abraham LincolnD'EverandTeam of Rivals: The Political Genius of Abraham LincolnÉvaluation : 4.5 sur 5 étoiles4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaD'EverandThe Unwinding: An Inner History of the New AmericaÉvaluation : 4 sur 5 étoiles4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreD'EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreÉvaluation : 4 sur 5 étoiles4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)D'EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Évaluation : 4.5 sur 5 étoiles4.5/5 (121)
- Her Body and Other Parties: StoriesD'EverandHer Body and Other Parties: StoriesÉvaluation : 4 sur 5 étoiles4/5 (821)
- Human Behavior and Social Environment: "The Individual Is A Biopsychosocial Being" Means That There IsDocument104 pagesHuman Behavior and Social Environment: "The Individual Is A Biopsychosocial Being" Means That There IsJerson GozarPas encore d'évaluation
- Unit I: Nursing Care of The Family Having Difficulty Conceiving A ChildDocument24 pagesUnit I: Nursing Care of The Family Having Difficulty Conceiving A ChildBlaise Anne InocPas encore d'évaluation
- Kandungan Protein, Lemak Daging Dan Kulit Itik, Entog Dan Mandalung Umur 8 MingguDocument5 pagesKandungan Protein, Lemak Daging Dan Kulit Itik, Entog Dan Mandalung Umur 8 MingguMaret MeiPas encore d'évaluation
- Liver Regeneration (PDFDrive)Document261 pagesLiver Regeneration (PDFDrive)Kapetan AhavPas encore d'évaluation
- Qt.2 Summative Test 1 & 2Document5 pagesQt.2 Summative Test 1 & 2Susan Salvador100% (1)
- AOSpine Manual Thoracic Spine AnatomyDocument18 pagesAOSpine Manual Thoracic Spine AnatomySebastián MorenoPas encore d'évaluation
- Acute MI DMII Medical ManagementDocument52 pagesAcute MI DMII Medical ManagementsherwincruzPas encore d'évaluation
- Blood ReportDocument17 pagesBlood ReportRajarajeshwari KommuPas encore d'évaluation
- Gut Dysfunction and Non-Alcoholic Fatty Liver DiseaseDocument9 pagesGut Dysfunction and Non-Alcoholic Fatty Liver DiseaseAshwiniPas encore d'évaluation
- Lipids, Biochem, ReviewerDocument9 pagesLipids, Biochem, ReviewerKerstine Anne AndradePas encore d'évaluation
- Endocrine System: Clarence L. Nuval, RMT, MDDocument66 pagesEndocrine System: Clarence L. Nuval, RMT, MDCla NuvalPas encore d'évaluation
- Scoles Pulst SCA2-Chapter 2018Document30 pagesScoles Pulst SCA2-Chapter 2018atpPas encore d'évaluation
- Angel Moon - Epilogue Haruka&MichiruDocument4 pagesAngel Moon - Epilogue Haruka&MichiruGarPas encore d'évaluation
- Medical Psychology Pidruch PDFDocument144 pagesMedical Psychology Pidruch PDFRaavanPas encore d'évaluation
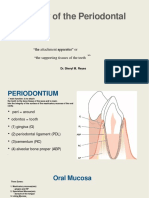
- THE PERIODONTIUM Copy 1Document98 pagesTHE PERIODONTIUM Copy 1Alexa AbrenicaPas encore d'évaluation
- Biology 2nd Lab Report (Onion Cell)Document4 pagesBiology 2nd Lab Report (Onion Cell)Johanna Haludilu88% (16)
- Mitosis and MeiosisDocument8 pagesMitosis and MeiosisShiela BelandresPas encore d'évaluation
- Digestion and Absorption of Carbohydrates: by Arpitha 3 BnysDocument31 pagesDigestion and Absorption of Carbohydrates: by Arpitha 3 BnysBig ZeroPas encore d'évaluation
- Analisis Kertas 3 Biologi SPMDocument2 pagesAnalisis Kertas 3 Biologi SPMsuhailinamohdikhwan100% (2)
- Date 01/dec/2021 04:33AM 30/nov/21 05:17PM Unit Bio Ref IntervalDocument2 pagesDate 01/dec/2021 04:33AM 30/nov/21 05:17PM Unit Bio Ref IntervalSaurabh PuriPas encore d'évaluation
- Movement of Particles NoteDocument2 pagesMovement of Particles NoteSabita SinghPas encore d'évaluation
- Basic Course Lectures in Ophthalmology 2013: Date Time Topic LecturerDocument2 pagesBasic Course Lectures in Ophthalmology 2013: Date Time Topic LecturerSteffi ClaroPas encore d'évaluation
- Pharmacology Practical (Dose Response Relationship of Carbachol and AtropineDocument11 pagesPharmacology Practical (Dose Response Relationship of Carbachol and AtropineESTHER WONG TZE YIING -Pas encore d'évaluation
- Blood PDFDocument4 pagesBlood PDFJustine May Balinggan DelmasPas encore d'évaluation
- Gills and Trachea. Moist Outer SkinDocument4 pagesGills and Trachea. Moist Outer SkinirisPas encore d'évaluation
- Answerd URT 2021Document8 pagesAnswerd URT 2021Abdo HusseinPas encore d'évaluation
- Detection and Identification of Antibodies - SCDocument67 pagesDetection and Identification of Antibodies - SCLyra Dennise LlidoPas encore d'évaluation
- Havening BookDocument124 pagesHavening BookSasa ZivkovicPas encore d'évaluation
- Week 2 - Enzyme Kinetics All - v2 - 3slide HandoutsDocument14 pagesWeek 2 - Enzyme Kinetics All - v2 - 3slide HandoutsFizza HussainPas encore d'évaluation
- Class 5 - Science Chapter 3Document18 pagesClass 5 - Science Chapter 3deviPas encore d'évaluation