Vous aimerez peut-être aussi
- Cours Complet Terminale-1 PDFDocument149 pagesCours Complet Terminale-1 PDFmamadou ndour100% (1)
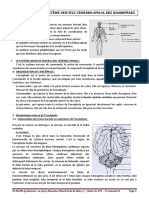
- Theme 1. Organisation Du Systeme Nerveux Cerebro-Spinal Des MammiferesDocument4 pagesTheme 1. Organisation Du Systeme Nerveux Cerebro-Spinal Des MammiferesFatou Diop100% (1)
- Cours Prof SVTDocument203 pagesCours Prof SVTAbibou Diop100% (1)
- Organisation de L'encéphaleDocument5 pagesOrganisation de L'encéphalesamuel BiléPas encore d'évaluation
- Le CerveauDocument10 pagesLe CerveauPonlue StephenPas encore d'évaluation
- Le CerveauDocument23 pagesLe CerveauksantiniPas encore d'évaluation
- Neuroanatomie Tome IIDocument78 pagesNeuroanatomie Tome IImohamed131983100% (1)
- Cours TS2 THIAWDocument117 pagesCours TS2 THIAWBaye ModouPas encore d'évaluation
- Les Tissus de Protection Du Syste0300me NerveuxDocument6 pagesLes Tissus de Protection Du Syste0300me NerveuxAmel YahiaPas encore d'évaluation
- 7AS1Document110 pages7AS1zeinebou sidi alhadiPas encore d'évaluation
- Le Tronc CérébralDocument5 pagesLe Tronc CérébralSaken667Pas encore d'évaluation
- ANATOMIE PR MejjatiDocument48 pagesANATOMIE PR MejjatiZakaria MaounPas encore d'évaluation
- Le Système CérébelleuxDocument4 pagesLe Système CérébelleuxMohammed GartitPas encore d'évaluation
- EncéphaleDocument9 pagesEncéphaleFéz EyPas encore d'évaluation
- Cours SVT TS-1-1Document127 pagesCours SVT TS-1-1Joseph Coly DioufPas encore d'évaluation
- Organisation de L'encéphaleDocument4 pagesOrganisation de L'encéphalePaul andre FayePas encore d'évaluation
- Chap 4 PHYSIO LENCEPHALEDocument5 pagesChap 4 PHYSIO LENCEPHALEAnani Jacques YaoPas encore d'évaluation
- 1 Anat CerveletDocument3 pages1 Anat Cerveletعمران عباسيPas encore d'évaluation
- Fascicule SVT TL2 2019-2020 ProfDocument62 pagesFascicule SVT TL2 2019-2020 Profbabacardiop2decPas encore d'évaluation
- Le Tronc CérébralDocument5 pagesLe Tronc Cérébraljoudjojo.06Pas encore d'évaluation
- Systeme NerveuxDocument82 pagesSysteme Nerveuxmohamedmahfoud198Pas encore d'évaluation
- Chapitre 6 Embryogenese Suite Neurulation DR Djeffal A1Document6 pagesChapitre 6 Embryogenese Suite Neurulation DR Djeffal A1Véto SamPas encore d'évaluation
- PDFDocument15 pagesPDFioana darkPas encore d'évaluation
- Anatomie CERVELETDocument3 pagesAnatomie CERVELETluxurystorino100% (1)
- Le Systeme NerveuxDocument4 pagesLe Systeme NerveuxCheymaPas encore d'évaluation
- Complément Ronéo Anatomie N°1Document5 pagesComplément Ronéo Anatomie N°1almnaouarPas encore d'évaluation
- GROUPE N°1, Embryologie Du Système Nerveux Central PDFDocument21 pagesGROUPE N°1, Embryologie Du Système Nerveux Central PDFMondePas encore d'évaluation
- Anatomie TOME IIIDocument32 pagesAnatomie TOME IIIHoUda ElGoPas encore d'évaluation
- Anatmie Externe Du CerveauxDocument8 pagesAnatmie Externe Du CerveauxMedecine DentairePas encore d'évaluation
- Neuro Anatomie ??Document170 pagesNeuro Anatomie ??Amirâ ZouânePas encore d'évaluation
- LCRDocument10 pagesLCRnaturopathe100% (1)
- Cours Histologie Du Cervelet 1Document11 pagesCours Histologie Du Cervelet 1Dany MelPas encore d'évaluation
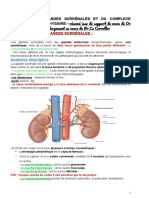
- GLANDES SURRÉNALES22-23-Version LibreDocument7 pagesGLANDES SURRÉNALES22-23-Version LibreLoubna KaouachiPas encore d'évaluation
- Activités L1 - L2Document3 pagesActivités L1 - L2Ahmadou KANDJIPas encore d'évaluation
- 1-Les Os Du CraneDocument15 pages1-Les Os Du Cranehocine benganaPas encore d'évaluation
- Neuro AnatDocument43 pagesNeuro AnatNouhaila AzirarPas encore d'évaluation
- Spé 12 - 10 - OreilleDocument5 pagesSpé 12 - 10 - OreillePASSLAS REUSSITEPas encore d'évaluation
- Le Tronc CérébralDocument4 pagesLe Tronc CérébralZaki RakiaPas encore d'évaluation
- Chap7 Le CerveletDocument2 pagesChap7 Le CerveletMahamane Hamissou Salissou Elhadji YahayaPas encore d'évaluation
- Anatomie de CCDocument12 pagesAnatomie de CCyoussef majdPas encore d'évaluation
- 3-Le Tronc CérébralDocument7 pages3-Le Tronc CérébralbPas encore d'évaluation
- 2 Generalites EmbryologiquesDocument6 pages2 Generalites EmbryologiquesHugues Roodly JeannitePas encore d'évaluation
- Systeme VentriculaireDocument46 pagesSysteme VentriculaireksantiniPas encore d'évaluation
- EncéphaleDocument40 pagesEncéphaleSamy Ta'y'b'iPas encore d'évaluation
- Eléments de Physiologie - Anatomie Du SNC Et Système Nerveux AutonomeDocument13 pagesEléments de Physiologie - Anatomie Du SNC Et Système Nerveux AutonomeMelPas encore d'évaluation
- Braillon Georges - Le Systeme Nerveux CentralDocument114 pagesBraillon Georges - Le Systeme Nerveux CentralMirela Visan0% (1)
- Système Nerveux DR BouraimaDocument61 pagesSystème Nerveux DR Bouraimaaba djibaPas encore d'évaluation
- TD - Système Nerveux - Sans TPDocument11 pagesTD - Système Nerveux - Sans TPFANKELE ALASSANE KONEPas encore d'évaluation
- Anatomie DIENCEPHALE-convertiDocument31 pagesAnatomie DIENCEPHALE-convertilordinondongo15Pas encore d'évaluation
- Chap 3 PHYSIOLOGIE DE LA MOELLE EPINIEREDocument13 pagesChap 3 PHYSIOLOGIE DE LA MOELLE EPINIEREAnani Jacques YaoPas encore d'évaluation
- Système NerveuxDocument11 pagesSystème Nerveuxhajar sektiouiPas encore d'évaluation
- La 4eme-8eme Semaine Cours 2018Document4 pagesLa 4eme-8eme Semaine Cours 2018130660LYC OMAR BENABDELAZIZ NEDROMAPas encore d'évaluation
- GastrulationDocument5 pagesGastrulationMinaheta Guy maurelPas encore d'évaluation
- Anatomie SNC Texte PDFDocument95 pagesAnatomie SNC Texte PDFJean PatatePas encore d'évaluation
- Chap3 Les Structures Pertinentes Du CerveauDocument3 pagesChap3 Les Structures Pertinentes Du CerveauMahamane Hamissou Salissou Elhadji YahayaPas encore d'évaluation
- 7 - MORPHOLOGIE DU TRONC CEREBRAL BlidaDocument12 pages7 - MORPHOLOGIE DU TRONC CEREBRAL BlidaRacha BznPas encore d'évaluation
- Fichier Produit 2272Document9 pagesFichier Produit 2272Amina HasnaouiPas encore d'évaluation
- Histologie Spe Üciale UCL Syst EndocrineDocument83 pagesHistologie Spe Üciale UCL Syst EndocrineJosué NAMEGABEPas encore d'évaluation
- Fiche Anatomie Nerf CranienDocument40 pagesFiche Anatomie Nerf Cranienemma.pailletPas encore d'évaluation
- Hémisphères cérébraux: Les Grands Articles d'UniversalisD'EverandHémisphères cérébraux: Les Grands Articles d'UniversalisPas encore d'évaluation
- 09 18-Catalogue-Single-Remedies - 21 09 18 - LRDocument20 pages09 18-Catalogue-Single-Remedies - 21 09 18 - LRAounAbdellahPas encore d'évaluation
- Tumeurs NeuroendocrinesDocument5 pagesTumeurs NeuroendocrinesAounAbdellahPas encore d'évaluation
- Theorie Homeo en 11 PointsDocument2 pagesTheorie Homeo en 11 PointsAounAbdellahPas encore d'évaluation
- Hyper VasopressinismeDocument1 pageHyper VasopressinismeAounAbdellah100% (1)
- Adénomes HypophysairesDocument8 pagesAdénomes HypophysairesAounAbdellahPas encore d'évaluation
- Néoplasies EndocriniennesDocument3 pagesNéoplasies EndocriniennesAounAbdellahPas encore d'évaluation
- Apopléxie AdénomateuseDocument1 pageApopléxie AdénomateuseAounAbdellahPas encore d'évaluation
- Syndrome Endocrinien de L'hémochromatose HéréditaireDocument2 pagesSyndrome Endocrinien de L'hémochromatose HéréditaireAounAbdellahPas encore d'évaluation
- Diabète InsipideDocument4 pagesDiabète InsipideAounAbdellahPas encore d'évaluation
- NutritionDocument2 pagesNutritionAounAbdellahPas encore d'évaluation
- AcromégalieDocument7 pagesAcromégalieAounAbdellahPas encore d'évaluation
- Q. VariéesDocument3 pagesQ. VariéesAounAbdellahPas encore d'évaluation
- Rein Et DiabèteDocument11 pagesRein Et DiabèteAounAbdellahPas encore d'évaluation
- ObésitéDocument1 pageObésitéAounAbdellahPas encore d'évaluation
- Neurofibromatose Type 1Document1 pageNeurofibromatose Type 1AounAbdellahPas encore d'évaluation
- Cancer de La ProstateDocument1 pageCancer de La ProstateAounAbdellahPas encore d'évaluation
- Les AnémiesDocument3 pagesLes AnémiesAounAbdellahPas encore d'évaluation
- Cancer Du SeinDocument1 pageCancer Du SeinAounAbdellahPas encore d'évaluation
- IRM CérébraleDocument1 pageIRM CérébraleAounAbdellahPas encore d'évaluation
- Echo-Cardiographie DopplerDocument1 pageEcho-Cardiographie DopplerAounAbdellahPas encore d'évaluation
- Rein Et MédicamentsDocument2 pagesRein Et MédicamentsAounAbdellahPas encore d'évaluation
- Sténoses Des Artères RénalesDocument2 pagesSténoses Des Artères RénalesAounAbdellahPas encore d'évaluation
- Décompensations Aigues Du DiabèreDocument5 pagesDécompensations Aigues Du DiabèreAounAbdellahPas encore d'évaluation
- TRT HypoglycémiantDocument6 pagesTRT HypoglycémiantAounAbdellahPas encore d'évaluation
- Syndrome MétaboliqueDocument3 pagesSyndrome MétaboliqueAounAbdellahPas encore d'évaluation
- DyslipidémiesDocument7 pagesDyslipidémiesAounAbdellahPas encore d'évaluation
- Diététique 1Document6 pagesDiététique 1AounAbdellahPas encore d'évaluation
- Relevé D'automesure TensionnelleDocument1 pageRelevé D'automesure TensionnelleAounAbdellahPas encore d'évaluation
- Diabète Et GrossesseDocument8 pagesDiabète Et GrossesseAounAbdellahPas encore d'évaluation
- ImprimerDocument9 pagesImprimerSawssan BstPas encore d'évaluation
- Anatomie UE5 Avec Codes CouleursDocument4 pagesAnatomie UE5 Avec Codes CouleursKate BlablaPas encore d'évaluation
- Physiologieam 150925134333 Lva1 App6892Document8 pagesPhysiologieam 150925134333 Lva1 App6892Mayssa BourenanePas encore d'évaluation
- Anatomie Radiologique de La Base Du Crâne Et Des Espaces Profonds de La FaceDocument68 pagesAnatomie Radiologique de La Base Du Crâne Et Des Espaces Profonds de La FaceNicu IacobPas encore d'évaluation
- Muscles MasticateursDocument25 pagesMuscles MasticateursMaram TanazeftiPas encore d'évaluation
- Anatomie-Physiologie-Locclusion DR Benazzouz 2022Document11 pagesAnatomie-Physiologie-Locclusion DR Benazzouz 2022riham fodhiliPas encore d'évaluation
- l3 Endocrino td1Document3 pagesl3 Endocrino td1Med MahmoudPas encore d'évaluation
- Résumé Des Nerfs CrâniensDocument16 pagesRésumé Des Nerfs Crâniensapi-3829498100% (1)
- TrachéeDocument33 pagesTrachéefatima nhariPas encore d'évaluation
- 047 Artères de La Cavité Nasale - Septum Nasal Relevé - Atlas Netter 19Document1 page047 Artères de La Cavité Nasale - Septum Nasal Relevé - Atlas Netter 19Ouma TaziPas encore d'évaluation
- QCM TP FinaleDocument18 pagesQCM TP FinaleReda AbouennajahPas encore d'évaluation
- Fichier Produit 2319Document45 pagesFichier Produit 2319Seydou KassoguePas encore d'évaluation
- Anatomie Du LarynxDocument8 pagesAnatomie Du LarynxPonlue StephenPas encore d'évaluation
- Anatomie 1Document52 pagesAnatomie 1Denisco NkemgnePas encore d'évaluation
- Voies Aériennes Supérieures (Cour)Document5 pagesVoies Aériennes Supérieures (Cour)Medjoudj SarahPas encore d'évaluation
- Thèse - Abakar OusmaneDocument80 pagesThèse - Abakar OusmaneThierno Madjou BAHPas encore d'évaluation
- Sebenta Neuroanatomia 2012 Volume IDocument96 pagesSebenta Neuroanatomia 2012 Volume IMariana VargasPas encore d'évaluation
- 4.le Système RespiratoireDocument5 pages4.le Système RespiratoireAndy Dilane EngoangPas encore d'évaluation
- Moelle Épinière OKDocument17 pagesMoelle Épinière OKMichel S. FayePas encore d'évaluation
- Corvilain 2018Document18 pagesCorvilain 2018Alex Martinez Jr.Pas encore d'évaluation
- Neurologie - Cours N°19 - Rousseau - Syndrome FrontalDocument18 pagesNeurologie - Cours N°19 - Rousseau - Syndrome FrontalCoralie PascardPas encore d'évaluation
- 2 - Physiologie PosturoDocDocument48 pages2 - Physiologie PosturoDocFrancisco Vicent PachecoPas encore d'évaluation
- 50538017Document285 pages50538017enzo BrdPas encore d'évaluation
- Recherches Associées: Structure Et Fonctionnement de L'oreill HumaineDocument1 pageRecherches Associées: Structure Et Fonctionnement de L'oreill HumaineCoralie PaysPas encore d'évaluation
- La Physiologie Du Systeme Nerveux Centrale DR GuedjatiDocument33 pagesLa Physiologie Du Systeme Nerveux Centrale DR GuedjatiMedMan6100% (2)
- Le Systeme NerveuxDocument3 pagesLe Systeme NerveuxoumPas encore d'évaluation
- 1 - Cours Neuropathologie Introduction Et Gliomes FMPR Anapath SpecialeDocument82 pages1 - Cours Neuropathologie Introduction Et Gliomes FMPR Anapath SpecialeMunir GuirouanPas encore d'évaluation
- Anatomie Topografica Testut Jacob 1921Document949 pagesAnatomie Topografica Testut Jacob 1921merac2000Pas encore d'évaluation
- Anatomie Tete Et Cou - ToppographiqueDocument41 pagesAnatomie Tete Et Cou - ToppographiquemzzdraPas encore d'évaluation