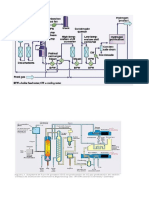
Vous aimerez peut-être aussi
- Inmovilizacion de CelulasDocument11 pagesInmovilizacion de CelulasFiorella Jennifer CZPas encore d'évaluation
- 13 Biotech Bioeng2004Document10 pages13 Biotech Bioeng2004Fiorella Jennifer CZPas encore d'évaluation
- ArtigoDocument6 pagesArtigoFelipe GomidePas encore d'évaluation
- Analisa KuantitatifDocument5 pagesAnalisa KuantitatifAswina NadiaPas encore d'évaluation
- Cellulase Production by Thermophilic Bacillus Sp. SMIA-2 and Its Detergent CompatibilityDocument10 pagesCellulase Production by Thermophilic Bacillus Sp. SMIA-2 and Its Detergent CompatibilitydadjPas encore d'évaluation
- Artigo DessalinationDocument6 pagesArtigo Dessalinationgswerne_335691290Pas encore d'évaluation
- Production of α-amylase using new strain of Bacillus polymyxa isolated from sweet potatoDocument7 pagesProduction of α-amylase using new strain of Bacillus polymyxa isolated from sweet potatoInternational Organization of Scientific Research (IOSR)Pas encore d'évaluation
- Bioresources.: Characteristics And Kinetic Analysis Of Β-Glucosidase (Mabgl) From Mucor Ardhlaengiktus Rsc1Document13 pagesBioresources.: Characteristics And Kinetic Analysis Of Β-Glucosidase (Mabgl) From Mucor Ardhlaengiktus Rsc1el batoPas encore d'évaluation
- Biotecnologia Vegetal. Cas 1 DefinitiuDocument10 pagesBiotecnologia Vegetal. Cas 1 DefinitiuAlbaPas encore d'évaluation
- Bacillus PolyfermenticusDocument6 pagesBacillus Polyfermenticussi275Pas encore d'évaluation
- Use of Uv-C Treatment For The Inactivation of MicroorganismsDocument6 pagesUse of Uv-C Treatment For The Inactivation of MicroorganismsGonzalo BoninoPas encore d'évaluation
- Purification y Characterization of Acidothermophilic Cellulase Enzyme Produced by Bacillus Subtilis Strain LF53 P09Document9 pagesPurification y Characterization of Acidothermophilic Cellulase Enzyme Produced by Bacillus Subtilis Strain LF53 P09Walter Michel Mamani MamaniPas encore d'évaluation
- Cultivation of Microalgae Chlorella Using Wine Industry By-ProductsDocument6 pagesCultivation of Microalgae Chlorella Using Wine Industry By-ProductsDANIELA YURLEY MENDOZA LIZCANOPas encore d'évaluation
- Alginte Beads FRMTNDocument8 pagesAlginte Beads FRMTNMeetika GuptaPas encore d'évaluation
- Production and Purification of Recombinant Human Interleukin-6 Secreted by The Yeast Saccharomyces CerevisiaeDocument6 pagesProduction and Purification of Recombinant Human Interleukin-6 Secreted by The Yeast Saccharomyces CerevisiaeNicolás Ignacio Valenzuela SayesPas encore d'évaluation
- Enzymatic Saccharification and Fermentation of CelDocument10 pagesEnzymatic Saccharification and Fermentation of CelSharmaPas encore d'évaluation
- The Influence of Lignin Degradation Products On Xylose Fermentation by Klebsiella PneumoniaeDocument4 pagesThe Influence of Lignin Degradation Products On Xylose Fermentation by Klebsiella Pneumoniaefilemon jaluPas encore d'évaluation
- Sdarticle 3Document6 pagesSdarticle 3nguyenthao146Pas encore d'évaluation
- Increased Xylitol Production Rate During Long-Term Cell Recycle Fermentation of Candida TropicalisDocument5 pagesIncreased Xylitol Production Rate During Long-Term Cell Recycle Fermentation of Candida TropicalisRAVI KUMARPas encore d'évaluation
- Carotenoid Production From Microalgae, Dunalilella SalinaDocument8 pagesCarotenoid Production From Microalgae, Dunalilella SalinaBernardo RodriguesPas encore d'évaluation
- Chen 1996Document4 pagesChen 1996mplatonosPas encore d'évaluation
- JF 1019104Document5 pagesJF 1019104Srotoswini JoardarPas encore d'évaluation
- Isolation and Purification of CellulaseDocument6 pagesIsolation and Purification of CellulaseMeshal NoorPas encore d'évaluation
- Asada y ShulerDocument7 pagesAsada y ShulerMARÍA DEL CARMEN NOVAL RAMÓNPas encore d'évaluation
- Enhanced Penicillin Production by Oligosaccharides From Batch Cultures of Penicillium Chrysogenum in Stirred-Tank ReactorsDocument6 pagesEnhanced Penicillin Production by Oligosaccharides From Batch Cultures of Penicillium Chrysogenum in Stirred-Tank Reactorsazzahra ghina fadillahPas encore d'évaluation
- Batra 2002Document6 pagesBatra 2002guiervicalPas encore d'évaluation
- β-galactosidase from Bacillus coagulans RCS3: Production and characterization of a thermostableDocument6 pagesβ-galactosidase from Bacillus coagulans RCS3: Production and characterization of a thermostablemoogambigai smPas encore d'évaluation
- Two-Stage Cultures For The Production of Astaxanthin From 6ialisDocument7 pagesTwo-Stage Cultures For The Production of Astaxanthin From 6ialisSaida ErradyPas encore d'évaluation
- 13 - 15v5i2 - 4 Serratia Marcescens OU50TDocument5 pages13 - 15v5i2 - 4 Serratia Marcescens OU50TIsworo RukmiPas encore d'évaluation
- TMP 633 EDocument5 pagesTMP 633 EFrontiersPas encore d'évaluation
- 15 PDFDocument7 pages15 PDFPradeep PereraPas encore d'évaluation
- Encapsulation of Lactobacillus CaseiDocument6 pagesEncapsulation of Lactobacillus CaseiAlejandro RebollarPas encore d'évaluation
- Decolourization of Azo Dye Methyl Red byDocument7 pagesDecolourization of Azo Dye Methyl Red byEvelyn NathaliaPas encore d'évaluation
- Immunostimulatory Principles From Chlorella Pyrenoidosa-Part 1: Isolation and Biological Assessment in VitroDocument8 pagesImmunostimulatory Principles From Chlorella Pyrenoidosa-Part 1: Isolation and Biological Assessment in VitroRunoffPas encore d'évaluation
- Role of Plasmid in Production of Acetobacter Xylinum BiofilmsDocument4 pagesRole of Plasmid in Production of Acetobacter Xylinum BiofilmsShofiaPas encore d'évaluation
- Nabarlatz2004 PDFDocument8 pagesNabarlatz2004 PDFAndrea PachonPas encore d'évaluation
- Antimicrobial Activity and Characteristics of Bacteriocins Produced by Vaginal LactobacilliDocument7 pagesAntimicrobial Activity and Characteristics of Bacteriocins Produced by Vaginal LactobacilliManohar PattarPas encore d'évaluation
- Art 01Document12 pagesArt 01Cristian-Catalin GavatPas encore d'évaluation
- (16187229 - E-Polymers) Hydrogel Beads From Sugar Cane Bagasse and Palm Kernel Cake, and The Viability of Encapsulated Lactobacillus AcidophilusDocument8 pages(16187229 - E-Polymers) Hydrogel Beads From Sugar Cane Bagasse and Palm Kernel Cake, and The Viability of Encapsulated Lactobacillus AcidophilusJuliana Cristina GomesPas encore d'évaluation
- Polysaccharides Produced by Two Red Algal Strains Grown in Digestate With Potential Ability of Tumor Cell InhibitionDocument11 pagesPolysaccharides Produced by Two Red Algal Strains Grown in Digestate With Potential Ability of Tumor Cell Inhibitionjuivanova7Pas encore d'évaluation
- Purification and Properties of A Keratinolytic Metalloprotease From Microbacterium SPDocument10 pagesPurification and Properties of A Keratinolytic Metalloprotease From Microbacterium SPFaisal AwanPas encore d'évaluation
- A New Method To Obtain B-Glucan From Saccharomyces Cerevisiae CellsDocument4 pagesA New Method To Obtain B-Glucan From Saccharomyces Cerevisiae Cellshuynhtanvu2011Pas encore d'évaluation
- Enantioselectivity of Sulcatone Reduction by Sorne Anaerobic BacteriaDocument8 pagesEnantioselectivity of Sulcatone Reduction by Sorne Anaerobic BacteriaFiorella Jennifer CZPas encore d'évaluation
- Solo 1Document9 pagesSolo 1JOSE MIGUEL ASMAD QUINTANAPas encore d'évaluation
- Ampling Techniques and ComparativeDocument6 pagesAmpling Techniques and ComparativeJessica Sanchez GutierrezPas encore d'évaluation
- Research Article: Optimization of Large-Scale Culture Conditions For The Production of Cordycepin WithDocument16 pagesResearch Article: Optimization of Large-Scale Culture Conditions For The Production of Cordycepin WithAsaddul IzzanPas encore d'évaluation
- 2008 - Selig et al - Heterologous Expression of Aspergillus niger β-d-XylosidaseDocument12 pages2008 - Selig et al - Heterologous Expression of Aspergillus niger β-d-XylosidaseRenata CalixtoPas encore d'évaluation
- Ardestani Et Al., 2015Document8 pagesArdestani Et Al., 201515041168Pas encore d'évaluation
- Purification and Partial Characterization of An Aldo-Keto Reductase From Saccharomyces CerevisiaeDocument6 pagesPurification and Partial Characterization of An Aldo-Keto Reductase From Saccharomyces CerevisiaeElmy Halily StudioPas encore d'évaluation
- Characterization of Industrially-Valuable Xylanopectinolytic Enzymes Produced Concurrently by A Novel Isolate of Bacillus PumilusDocument9 pagesCharacterization of Industrially-Valuable Xylanopectinolytic Enzymes Produced Concurrently by A Novel Isolate of Bacillus PumilusLiyin WongPas encore d'évaluation
- 2.2.1. Sample Collection: BT Isolation Was Done by Using Two Different Methods Based On The Sample Type. The ShakenDocument4 pages2.2.1. Sample Collection: BT Isolation Was Done by Using Two Different Methods Based On The Sample Type. The ShakenLeong HlPas encore d'évaluation
- Micro-Encapsulation of Bifidobacterium Lactis For Incorporation Into Soft FoodsDocument6 pagesMicro-Encapsulation of Bifidobacterium Lactis For Incorporation Into Soft FoodsArisbet SantisPas encore d'évaluation
- Rafael EjemplosDocument7 pagesRafael EjemplosAntonio MoncayoPas encore d'évaluation
- Isolation and Characterization of A Thermostable - Amylase FromDocument5 pagesIsolation and Characterization of A Thermostable - Amylase FromDương Nguyễn Thùy DungPas encore d'évaluation
- Production of Ethanol From MolassesDocument5 pagesProduction of Ethanol From MolassesKrishna PotluriPas encore d'évaluation
- 4928 18017 1 PB PDFDocument8 pages4928 18017 1 PB PDFErik Javier Jefe MicrobiologiaPas encore d'évaluation
- Uquiche 2019Document8 pagesUquiche 2019Ommi Samuel G SPas encore d'évaluation
- Phellinus Linteus: Structural and Biological Study of Carboxymethylated PolysaccharidesDocument5 pagesPhellinus Linteus: Structural and Biological Study of Carboxymethylated PolysaccharidesMinh NguyễnPas encore d'évaluation
- Pan, X., Chen, F., Wu, T., Tang, H., Dan Zhao, Z. 2009. The Acid, Bile Tolerance and Antimicrobial Property of Lactobacillus Acidophilus NIT. J. Food Control. 20 598-602.Document5 pagesPan, X., Chen, F., Wu, T., Tang, H., Dan Zhao, Z. 2009. The Acid, Bile Tolerance and Antimicrobial Property of Lactobacillus Acidophilus NIT. J. Food Control. 20 598-602.AfdhalRuslanPas encore d'évaluation
- Enantioselectivity of Sulcatone Reduction by Sorne Anaerobic BacteriaDocument8 pagesEnantioselectivity of Sulcatone Reduction by Sorne Anaerobic BacteriaFiorella Jennifer CZPas encore d'évaluation
- Brad Pitt BibliographyDocument1 pageBrad Pitt BibliographyFiorella Jennifer CZPas encore d'évaluation
- T (S) V X 106 (m3) V (cm3) V/A (CM) At/V (S/CM)Document2 pagesT (S) V X 106 (m3) V (cm3) V/A (CM) At/V (S/CM)Fiorella Jennifer CZPas encore d'évaluation
- Filtración de Cristalesde Gentamicina: T (Sec) V (L) T/VDocument2 pagesFiltración de Cristalesde Gentamicina: T (Sec) V (L) T/VFiorella Jennifer CZPas encore d'évaluation
- Speaking 2 Intensive 6Document1 pageSpeaking 2 Intensive 6Fiorella Jennifer CZPas encore d'évaluation
- Diagram AsDocument2 pagesDiagram AsFiorella Jennifer CZPas encore d'évaluation
- Bio Problem 2Document8 pagesBio Problem 2Fiorella Jennifer CZPas encore d'évaluation
- Speaking 1Document1 pageSpeaking 1Fiorella Jennifer CZPas encore d'évaluation
- FI G. 1: Refri Geraci Ón Por C Om Presi Ón - L I Cuefa Cci ÓnDocument3 pagesFI G. 1: Refri Geraci Ón Por C Om Presi Ón - L I Cuefa Cci ÓnFiorella Jennifer CZPas encore d'évaluation
- Pio 2Document2 pagesPio 2Fiorella Jennifer CZPas encore d'évaluation
- Speaking 1Document1 pageSpeaking 1Fiorella Jennifer CZPas encore d'évaluation
- S and P Block ElementsDocument47 pagesS and P Block ElementsDhruv SainiPas encore d'évaluation
- GUID - 4 en-USDocument2 pagesGUID - 4 en-USEileen Mae ChanPas encore d'évaluation
- New Expanders For New Battery Applications. Start/Stop and HEVDocument31 pagesNew Expanders For New Battery Applications. Start/Stop and HEVPaul OñaPas encore d'évaluation
- Fulltext01 13Document64 pagesFulltext01 13sheberuaPas encore d'évaluation
- Paper - A Review For Characterization of Silicafume & Its Effect On ConcreteDocument8 pagesPaper - A Review For Characterization of Silicafume & Its Effect On ConcreteAnto Destianto100% (1)
- Name Raj Goswami: Course Name Class Year Subject Code Subject Name InstituteDocument25 pagesName Raj Goswami: Course Name Class Year Subject Code Subject Name InstituteRaj GoswamiPas encore d'évaluation
- Durebild STE: Surface Tolerant High Build High Solids Epoxy CoatingDocument3 pagesDurebild STE: Surface Tolerant High Build High Solids Epoxy CoatinglivefreakPas encore d'évaluation
- Bu 2022Document10 pagesBu 2022Rafael RodriguesPas encore d'évaluation
- 2016 1113 Oct P1Document9 pages2016 1113 Oct P1Duong Anh HoàngPas encore d'évaluation
- EI-014 Rev.1 - GroutDocument11 pagesEI-014 Rev.1 - GroutDavid Ricardo Lozano RondonPas encore d'évaluation
- Stoichiometry The Accounting in ChemistryDocument28 pagesStoichiometry The Accounting in ChemistrySamKris Guerrero MalasagaPas encore d'évaluation
- Mutu International - Laboratory Profile ALL - 20.9 2022Document40 pagesMutu International - Laboratory Profile ALL - 20.9 2022Iwan FauziPas encore d'évaluation
- Inorganic Chemistry: Electron ConfigurationDocument2 pagesInorganic Chemistry: Electron ConfigurationArah LlamasPas encore d'évaluation
- Pneumatic Conveying SystemDocument27 pagesPneumatic Conveying SystemRavindra DarjiPas encore d'évaluation
- 5Document72 pages5Hasna RedPas encore d'évaluation
- DSC Analysis of Plastics FinalDocument3 pagesDSC Analysis of Plastics FinalFan ServicesNSKPas encore d'évaluation
- D NA Vs RNA: A Comparison in Structure and FunctionDocument3 pagesD NA Vs RNA: A Comparison in Structure and FunctionHossam GamilPas encore d'évaluation
- 2.4 Basic Concepts of Crystal StructureDocument7 pages2.4 Basic Concepts of Crystal Structurecory kurdapyaPas encore d'évaluation
- ISO 7253 2001 Method Statement PDFDocument4 pagesISO 7253 2001 Method Statement PDFFadi MagdyPas encore d'évaluation
- Periodic Properties of The Elements 608817Document3 pagesPeriodic Properties of The Elements 608817Kumar nayakPas encore d'évaluation
- Keraghel 2020Document16 pagesKeraghel 2020dj SidPas encore d'évaluation
- Question Bank The Periodic Table: Reasons?Document13 pagesQuestion Bank The Periodic Table: Reasons?Mehul MayankPas encore d'évaluation
- CHEM110-Spring 2021 - 2022-Experiment 3Document14 pagesCHEM110-Spring 2021 - 2022-Experiment 3bm7gyygjtfPas encore d'évaluation
- Imdg Segregation Table: Refer To IMDG Code and Relevant Marine NoticesDocument1 pageImdg Segregation Table: Refer To IMDG Code and Relevant Marine NoticesIain RobbPas encore d'évaluation
- CO + H O H + Hco: Overall ReactionDocument12 pagesCO + H O H + Hco: Overall ReactionLuke ShantiPas encore d'évaluation
- Perforation: Omega 2013Document73 pagesPerforation: Omega 2013Igbereyivwe TejiriPas encore d'évaluation
- ASNT NDT Level II ExaminationDocument11 pagesASNT NDT Level II ExaminationPardie YanPas encore d'évaluation
- Feasibility Study of Invert Sugar ProductionDocument3 pagesFeasibility Study of Invert Sugar ProductionIntratec SolutionsPas encore d'évaluation
- Cell and Molecular BiologyDocument18 pagesCell and Molecular BiologyHizzei Caballero100% (1)
- Mole Concept A 4Document57 pagesMole Concept A 4rakeshpoddarPas encore d'évaluation