Vous aimerez peut-être aussi
- Research Article: Composition of Algal Oil and Its Potential As BiofuelDocument15 pagesResearch Article: Composition of Algal Oil and Its Potential As BiofuelTeresa MataPas encore d'évaluation
- Review Article: Algal Biofuels: A Credible Prospective?Document15 pagesReview Article: Algal Biofuels: A Credible Prospective?BASKAR EPas encore d'évaluation
- Solar Hydrogen Energy 2Document38 pagesSolar Hydrogen Energy 2ignacioPas encore d'évaluation
- Biodiesel From Algae: Challenges and ProspectsDocument10 pagesBiodiesel From Algae: Challenges and ProspectsRahul KumarPas encore d'évaluation
- Progress in Nuclear Energy: Charles W. ForsbergDocument9 pagesProgress in Nuclear Energy: Charles W. ForsbergJosé Henrique Holtz PeovezanPas encore d'évaluation
- Applied Energy: Amita Jacob, Ao Xia, Jerry D. MurphyDocument7 pagesApplied Energy: Amita Jacob, Ao Xia, Jerry D. MurphyAgustine Christela MelvianaPas encore d'évaluation
- Designing of a PV/Wind/Diesel Hybrid Energy SystemD'EverandDesigning of a PV/Wind/Diesel Hybrid Energy SystemÉvaluation : 5 sur 5 étoiles5/5 (2)
- Algae Oil EconomicsDocument12 pagesAlgae Oil EconomicsDfox123456Pas encore d'évaluation
- CO2 Sequestration by Algae Reactors: Calling It Unproved Technology Understates Its ProblemsDocument1 pageCO2 Sequestration by Algae Reactors: Calling It Unproved Technology Understates Its ProblemsanuanuaPas encore d'évaluation
- Research PaperDocument9 pagesResearch PaperCody RickmanPas encore d'évaluation
- Biofuels From Microalgae PDFDocument6 pagesBiofuels From Microalgae PDFluis APPas encore d'évaluation
- BiomassDocument24 pagesBiomassnasir608100% (1)
- Literature Review On Petroleum ProductsDocument8 pagesLiterature Review On Petroleum Productsafdtunqho100% (1)
- 1-s2.0-S1364032115003998-Yanwen Shen Renewable Fuel 2015Document52 pages1-s2.0-S1364032115003998-Yanwen Shen Renewable Fuel 2015Rahmi ArslanPas encore d'évaluation
- Environmental Aspects Oof Energy UtilizationDocument3 pagesEnvironmental Aspects Oof Energy UtilizationHemanth Raju0% (1)
- Estimation of Liquid Fuel Yields From Biomass: Environ. Sci. Technol. 2010, 44, 5298-5305Document8 pagesEstimation of Liquid Fuel Yields From Biomass: Environ. Sci. Technol. 2010, 44, 5298-5305yemresimsekPas encore d'évaluation
- 6056f41af339e4-Powering Offshore Structure Using Renewable EnergyDocument20 pages6056f41af339e4-Powering Offshore Structure Using Renewable EnergyRadhin Hanif FathurrahmanPas encore d'évaluation
- Kunj Apur 2010Document11 pagesKunj Apur 2010Jamie SamuelPas encore d'évaluation
- Photobioreactor Design For Commercial Biofuel Production From Microalgae PDFDocument11 pagesPhotobioreactor Design For Commercial Biofuel Production From Microalgae PDFBrei Parayno LaurioPas encore d'évaluation
- Tutorial Renewable EnergyDocument26 pagesTutorial Renewable Energyak alifPas encore d'évaluation
- Are Biofuels The Best Use of SunlightDocument27 pagesAre Biofuels The Best Use of SunlightSureshPas encore d'évaluation
- MARSTEC 08 - CAMERA READY PAPER - Environmetal Risk Based Design For Waste Derived Bio-EnergyDocument13 pagesMARSTEC 08 - CAMERA READY PAPER - Environmetal Risk Based Design For Waste Derived Bio-EnergyoladokunsulaimanPas encore d'évaluation
- Fuels. Fuels Fuels: Stern, 2006Document7 pagesFuels. Fuels Fuels: Stern, 2006annjie99Pas encore d'évaluation
- Microalgae: The Potential For Carbon Capture: Articles ArticlesDocument6 pagesMicroalgae: The Potential For Carbon Capture: Articles ArticlesCristian SotoPas encore d'évaluation
- Combining Solar Photovoltaic Panels and Food Crops For Optimising Land UseDocument8 pagesCombining Solar Photovoltaic Panels and Food Crops For Optimising Land UseJULIANAPas encore d'évaluation
- A Project On Solar Battery ChargerDocument27 pagesA Project On Solar Battery ChargerVaibhav Charanti89% (53)
- De La Fotosíntesis Natural A La ArtificialDocument16 pagesDe La Fotosíntesis Natural A La ArtificialHugo Nieto BarreraPas encore d'évaluation
- Chapter 1 IntroductionDocument6 pagesChapter 1 Introductionسالم شعيبPas encore d'évaluation
- Renewable and Non-Renewable ResourcesDocument12 pagesRenewable and Non-Renewable ResourceschetanPas encore d'évaluation
- Biohydrogen Production: Prospectsand Limitationsto Practical ApplicationDocument13 pagesBiohydrogen Production: Prospectsand Limitationsto Practical ApplicationFirman teguh santosoPas encore d'évaluation
- Solar Energy 2008Document58 pagesSolar Energy 2008Prakash VenkatesanPas encore d'évaluation
- Biophysical Chemistry: Diana Simionato, Stefania Basso, Giorgio M. Giacometti, Tomas MorosinottoDocument8 pagesBiophysical Chemistry: Diana Simionato, Stefania Basso, Giorgio M. Giacometti, Tomas MorosinottoAngel Quijano ArmengolPas encore d'évaluation
- Analysis of A Waste-To-Energy Power Plant With CO SequestrationDocument8 pagesAnalysis of A Waste-To-Energy Power Plant With CO SequestrationFerdiansyah Iqbal RafandiPas encore d'évaluation
- Algae FuelDocument5 pagesAlgae Fuelphoebus101Pas encore d'évaluation
- Ii.A.2. Are There Biogeophysical Limits To Growth?: Scenarios For The 21 CenturyDocument32 pagesIi.A.2. Are There Biogeophysical Limits To Growth?: Scenarios For The 21 Centuryapi-51069617Pas encore d'évaluation
- Lecture 1Document20 pagesLecture 1Sparsh ThakurPas encore d'évaluation
- Article 2 ADocument13 pagesArticle 2 AFJosue MalaveHPas encore d'évaluation
- Energy Production From Biomass (Part 1)Document10 pagesEnergy Production From Biomass (Part 1)36turkPas encore d'évaluation
- Merged Document 2Document90 pagesMerged Document 2Anonymous flQkUjZCPas encore d'évaluation
- ReportDocument47 pagesReportmayank911Pas encore d'évaluation
- A Study On The Potential of Corn Cob Enginegenerator For ElectriDocument4 pagesA Study On The Potential of Corn Cob Enginegenerator For ElectriCeddie MianoPas encore d'évaluation
- 9 Principles of Photobioreactor DesignDocument30 pages9 Principles of Photobioreactor DesignNadinePas encore d'évaluation
- Biorefinery - The Worldwide Status at The Beginning of 2006Document62 pagesBiorefinery - The Worldwide Status at The Beginning of 2006José Augusto Travassos ToméPas encore d'évaluation
- Nuclear Energy in the 21st Century: World Nuclear University PressD'EverandNuclear Energy in the 21st Century: World Nuclear University PressÉvaluation : 4.5 sur 5 étoiles4.5/5 (3)
- Chapter 4Document24 pagesChapter 4Manuel Alejandro Estrella GutiérrezPas encore d'évaluation
- Biotechnology Advances: Niels-Henrik Norsker, Maria J. Barbosa, Marian H. Vermuë, René H. WijffelsDocument4 pagesBiotechnology Advances: Niels-Henrik Norsker, Maria J. Barbosa, Marian H. Vermuë, René H. WijffelsEng MohammedPas encore d'évaluation
- Biomass Conversion PDFDocument8 pagesBiomass Conversion PDFVivek Rajkumar GPas encore d'évaluation
- Grow AlgaeDocument6 pagesGrow AlgaeravindrawariPas encore d'évaluation
- Heading Passage 5Document3 pagesHeading Passage 5Vân Khánh TrầnPas encore d'évaluation
- A Blueprint For Achieving Net-Zero CO2 Emissions: The Difficult Road for the US to Achieve Net-Zero CO2 Emissions by 2050D'EverandA Blueprint For Achieving Net-Zero CO2 Emissions: The Difficult Road for the US to Achieve Net-Zero CO2 Emissions by 2050Pas encore d'évaluation
- Algae To Energy - PowerpointDocument24 pagesAlgae To Energy - PowerpointjkhrashprashPas encore d'évaluation
- 1 s2.0 S1364032120305670 MainDocument15 pages1 s2.0 S1364032120305670 MainJhansi VaranasiPas encore d'évaluation
- Importance of Nuclear EnergyDocument91 pagesImportance of Nuclear EnergyMuhammad NasirPas encore d'évaluation
- Hydrogen Economy Based On Renewable EnerDocument8 pagesHydrogen Economy Based On Renewable EnerPanche AtanasovskiPas encore d'évaluation
- Renewable Energy: Name: Ahmed Mohamed Farouk Abdo Reg: 08100954Document11 pagesRenewable Energy: Name: Ahmed Mohamed Farouk Abdo Reg: 08100954Ahab CaptainPas encore d'évaluation
- Cultivating Liquid Algae For Fuel ProductionDocument6 pagesCultivating Liquid Algae For Fuel ProductionNeil100% (3)
- Endoscopic Retrival DevicesDocument6 pagesEndoscopic Retrival Deviceschennai1stPas encore d'évaluation
- Mushroom Economics in ChinaDocument4 pagesMushroom Economics in Chinachennai1stPas encore d'évaluation
- Glycemic Index LoadDocument52 pagesGlycemic Index Loadchennai1stPas encore d'évaluation
- Soilless Gardening 01Document52 pagesSoilless Gardening 01Naveen Kumar GuptaPas encore d'évaluation
- NAEP vs. TIMSS vs. PISA ComparisonDocument11 pagesNAEP vs. TIMSS vs. PISA Comparisonchennai1stPas encore d'évaluation
- Greenhouse Management and Enterprise BrochureDocument2 pagesGreenhouse Management and Enterprise Brochurechennai1stPas encore d'évaluation
- Subsurface Textile IrrigationDocument2 pagesSubsurface Textile Irrigationchennai1st50% (2)
- Pies and Plates Healthy Living Kitchen - MushroomsDocument14 pagesPies and Plates Healthy Living Kitchen - Mushroomschennai1stPas encore d'évaluation
- Lessons To Be Learned From The Finnish School SystemDocument13 pagesLessons To Be Learned From The Finnish School Systemchennai1stPas encore d'évaluation
- Yoga For Women ArticleDocument8 pagesYoga For Women ArticleVasavi JalapatiPas encore d'évaluation
- Tamil ProverbsDocument534 pagesTamil Proverbschennai1st100% (10)
- Temperature For Growing StrawberriesDocument9 pagesTemperature For Growing Strawberrieschennai1stPas encore d'évaluation
- Greenhouse Vegetable Production Economic ConsiderationsDocument15 pagesGreenhouse Vegetable Production Economic Considerationschennai1stPas encore d'évaluation
- The Production of CompostDocument7 pagesThe Production of Compostchennai1stPas encore d'évaluation
- Banana CultivationDocument33 pagesBanana Cultivationshashankdatey100% (2)
- Fruit and Vegetable Waste Hydrolysates As Growth Medium For Higher Biomass and Lipid Production in Chlorella VulgarisDocument7 pagesFruit and Vegetable Waste Hydrolysates As Growth Medium For Higher Biomass and Lipid Production in Chlorella VulgarisPremier PublishersPas encore d'évaluation
- Utilization of Microalgal Bacterial Energy Nexus Improves - 2023 - EcotoxicologyDocument16 pagesUtilization of Microalgal Bacterial Energy Nexus Improves - 2023 - EcotoxicologyCristhian RodriguezPas encore d'évaluation
- Mass Balance For Biomass and Biogas Production From Organic WasteDocument14 pagesMass Balance For Biomass and Biogas Production From Organic WastedavidchemwenoPas encore d'évaluation
- NAME - Sahil Yadav Class - Xiith Roll NODocument60 pagesNAME - Sahil Yadav Class - Xiith Roll NOAnkit YadavPas encore d'évaluation
- Generation and Harvesting of Microalgae Biomass For Biofuel ProductionDocument23 pagesGeneration and Harvesting of Microalgae Biomass For Biofuel ProductionFábio UcellaPas encore d'évaluation
- Chapter 7Document14 pagesChapter 7Elena Rojo de BenitoPas encore d'évaluation
- 8 - Lucas PedevillaDocument16 pages8 - Lucas Pedevillap706e01Pas encore d'évaluation
- White Paper On Sustainable Jet Fuel June 2012 Faaij Van DijkDocument18 pagesWhite Paper On Sustainable Jet Fuel June 2012 Faaij Van DijkMax RomeroPas encore d'évaluation
- Extraction of Oil From Algae For Biodiesel ProductDocument8 pagesExtraction of Oil From Algae For Biodiesel ProductAbdisa GemechuPas encore d'évaluation
- Determination of Total Lipids As Fatty Acid Methyl Esters (FAME) by in Situ TransesterificationDocument16 pagesDetermination of Total Lipids As Fatty Acid Methyl Esters (FAME) by in Situ TransesterificationpeterPas encore d'évaluation
- Microalgae An Ecofriendly and Sustainable Wastewater Treatment OptionDocument10 pagesMicroalgae An Ecofriendly and Sustainable Wastewater Treatment OptionEustache NIJEJEPas encore d'évaluation
- Bio-Fuel IndonesiaDocument26 pagesBio-Fuel IndonesiatcandelarioPas encore d'évaluation
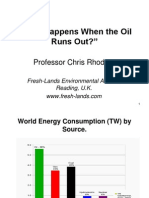
- Chris Rhodes - What Happens When The Oil Runs OutDocument44 pagesChris Rhodes - What Happens When The Oil Runs OutOnTheWightNewsPas encore d'évaluation
- Chlorella For Protein and Biofuels: From Strain Selection To Outdoor Cultivation in A Green Wall Panel PhotobioreactorDocument12 pagesChlorella For Protein and Biofuels: From Strain Selection To Outdoor Cultivation in A Green Wall Panel PhotobioreactorEssam KamalPas encore d'évaluation
- Algae 2020Document4 pagesAlgae 2020Samran AliPas encore d'évaluation
- Biofuel For Sri LankaDocument0 pageBiofuel For Sri LankaJaanu SanthiranPas encore d'évaluation
- Electricity From AlgaeDocument9 pagesElectricity From AlgaeRaghamath KaniPas encore d'évaluation
- Energies 05 00243Document15 pagesEnergies 05 00243Adam WilsonPas encore d'évaluation
- Biomass and Biofuels From Microalgae (Navid)Document373 pagesBiomass and Biofuels From Microalgae (Navid)Eu Be Rahat100% (1)
- Biodiesel From Micro AlgaeDocument3 pagesBiodiesel From Micro Algaeraanja2Pas encore d'évaluation
- Sustainable Bio Kerosene: Process Routes and Industrial Demonstration Activities in Aviation BiofuelsDocument9 pagesSustainable Bio Kerosene: Process Routes and Industrial Demonstration Activities in Aviation BiofuelsJaymar JaminPas encore d'évaluation
- DRC Presentation Revised - MS FinalDocument18 pagesDRC Presentation Revised - MS FinalDr Monika SoganiPas encore d'évaluation
- Ibr Arra SolazymeDocument1 pageIbr Arra SolazymeRamakrishnan TamilvaananPas encore d'évaluation
- Ethanol From Sugar Beets - A Process and Economic AnalysisDocument143 pagesEthanol From Sugar Beets - A Process and Economic Analysisgigi7980Pas encore d'évaluation
- PHOTOBIOREACTORDocument17 pagesPHOTOBIOREACTORParmeshwarPaulPas encore d'évaluation
- 12 - Aviation and The EnvironmentDocument30 pages12 - Aviation and The EnvironmentJurandir SanchesPas encore d'évaluation
- Egfi Engineering Go For It MagazineDocument81 pagesEgfi Engineering Go For It Magazineapi-243633061Pas encore d'évaluation
- Designing Controls For The Process Industries 1st Seames Solution ManualDocument6 pagesDesigning Controls For The Process Industries 1st Seames Solution ManualElizabethVancejodf100% (43)
- Bio ProjectDocument17 pagesBio Projectsaifnkhannrahman100% (5)
- Prospects of Biodiesel Production From Microalgae in IndiaDocument12 pagesProspects of Biodiesel Production From Microalgae in IndiaAditya KarpePas encore d'évaluation