Vous aimerez peut-être aussi
- Lead Funnels On Funnel Swipe File - TrelloDocument5 pagesLead Funnels On Funnel Swipe File - TrelloKatherie BriersPas encore d'évaluation
- List of Electronics Manufacturing Companies in Noida - ElectronicsmediaDocument2 pagesList of Electronics Manufacturing Companies in Noida - ElectronicsmediaBlue Oceon50% (4)
- Molecular Cell Biology Lodish 7th Edition Solutions ManualDocument8 pagesMolecular Cell Biology Lodish 7th Edition Solutions ManualVictoriaWilliamsegtnm100% (82)
- 2 Acceleration Questions and AnswersDocument2 pages2 Acceleration Questions and Answersapi-272986951100% (2)
- World MultipleDocument271 pagesWorld MultipleatrkpolePas encore d'évaluation
- Cervical CellDocument9 pagesCervical Cellsakata_abera4Pas encore d'évaluation
- Methods For Monitoring Cellular Motion and FunctionDocument4 pagesMethods For Monitoring Cellular Motion and FunctionDiogo MoreiraPas encore d'évaluation
- Geometric Approach To Segmentation and Protein Localization in Cell Cultured AssaysDocument10 pagesGeometric Approach To Segmentation and Protein Localization in Cell Cultured AssaysglivanosPas encore d'évaluation
- Improved Automatic Detection and Segmentation of Cell Nuclei in Histopathology ImagesDocument12 pagesImproved Automatic Detection and Segmentation of Cell Nuclei in Histopathology ImagesEgor FenkoPas encore d'évaluation
- Flow cytometry analysis and cell sorting in plants: past developments and future potentialDocument6 pagesFlow cytometry analysis and cell sorting in plants: past developments and future potentialTallie ZeidlerPas encore d'évaluation
- A Deep Learning-Based Algorithm For 2-D Cell Segmentation in Microscopy ImagesDocument11 pagesA Deep Learning-Based Algorithm For 2-D Cell Segmentation in Microscopy ImagesIgnasi AlemanyPas encore d'évaluation
- Chapter 3: Analysis of Dendritic Arborization and Spine MorphologyDocument14 pagesChapter 3: Analysis of Dendritic Arborization and Spine MorphologyAshish KumarPas encore d'évaluation
- Label-Free Imaging of Membrane Potential Using Membrane ElectromotilityDocument8 pagesLabel-Free Imaging of Membrane Potential Using Membrane ElectromotilityNur AnindyaPas encore d'évaluation
- NatCommBucher2018 PDFDocument13 pagesNatCommBucher2018 PDFRichiePas encore d'évaluation
- Software Defect Identification Using MacDocument10 pagesSoftware Defect Identification Using Macwmarasigan2610Pas encore d'évaluation
- Cell Identificación and Sizing Using Digital Image Analysis For Estimation of Cell Biomass in High Rate Algal PondsDocument12 pagesCell Identificación and Sizing Using Digital Image Analysis For Estimation of Cell Biomass in High Rate Algal PondsAldemar CortesPas encore d'évaluation
- Article Conférence ADRAR 2017Document5 pagesArticle Conférence ADRAR 2017Benomar AminePas encore d'évaluation
- Research Article: The Café Wall Illusion: Local and Global Perception From Multiple Scales To MultiscaleDocument23 pagesResearch Article: The Café Wall Illusion: Local and Global Perception From Multiple Scales To MultiscaleLas UkcuPas encore d'évaluation
- Computers in Biology and Medicine: B. Nayebifar, H. Abrishami MoghaddamDocument8 pagesComputers in Biology and Medicine: B. Nayebifar, H. Abrishami MoghaddamMohammad RofiiPas encore d'évaluation
- Estimating The Size of Onion Epidermal Cells From Diffraction PatternsDocument5 pagesEstimating The Size of Onion Epidermal Cells From Diffraction PatternsAlexis Nathán RuedaPas encore d'évaluation
- 2011 Meijering Et Al Particle Tracking MethodDocument18 pages2011 Meijering Et Al Particle Tracking Methodhansraj4u4800Pas encore d'évaluation
- Cell Adhesion StudyDocument11 pagesCell Adhesion StudyYashaswy GovadaPas encore d'évaluation
- Blood Vessel Segmentation in Fundus ImagesDocument5 pagesBlood Vessel Segmentation in Fundus ImagesNaoufal NaoufalPas encore d'évaluation
- Studies in Molecular Dynamics. II. Behavior of A Small Number of Elastic SpheresDocument14 pagesStudies in Molecular Dynamics. II. Behavior of A Small Number of Elastic SpheresJustin BrockPas encore d'évaluation
- Liver Segmentation in CT Images Using Three Dimensional To Two Dimensional Fully Convolutional NetworkDocument5 pagesLiver Segmentation in CT Images Using Three Dimensional To Two Dimensional Fully Convolutional Networkmanju.dsatmPas encore d'évaluation
- Thesis Office UiucDocument5 pagesThesis Office Uiucbopajagenal2100% (2)
- Department of Physics College of Natural and Computational Sciences Addis Ababa UniversityDocument11 pagesDepartment of Physics College of Natural and Computational Sciences Addis Ababa UniversityHenock TadelePas encore d'évaluation
- TMP 51 BDocument7 pagesTMP 51 BFrontiersPas encore d'évaluation
- O. Tauri: 3-D Ultrastructure Of: Electron Cryotomography of An Entire Eukaryotic CellDocument14 pagesO. Tauri: 3-D Ultrastructure Of: Electron Cryotomography of An Entire Eukaryotic CellMARIA ANGGIE CANTIKA DEWANIPas encore d'évaluation
- Processing of DNA and Protein Electrophoresis Gels by Image AnalysisDocument8 pagesProcessing of DNA and Protein Electrophoresis Gels by Image AnalysisnurhidayahsidikPas encore d'évaluation
- 1Document13 pages1tanjiromanjiro20Pas encore d'évaluation
- Real-Time Volumetric Microscopy of in Vivo Dynamics and Large-Scale Samples With SCAPE 2.0Document15 pagesReal-Time Volumetric Microscopy of in Vivo Dynamics and Large-Scale Samples With SCAPE 2.0Honghai LiuPas encore d'évaluation
- Transmission Electron Microscopy Research PapersDocument4 pagesTransmission Electron Microscopy Research PapersgvzexnzhPas encore d'évaluation
- Erythrocyte Structure and Dynamics Quantified by Hilbert Phase MicrosDocument3 pagesErythrocyte Structure and Dynamics Quantified by Hilbert Phase MicrosSendyka RinduwastutyPas encore d'évaluation
- High-Speed Fluorescence Image - Enabled Cell Sorting: Research TechnologyDocument7 pagesHigh-Speed Fluorescence Image - Enabled Cell Sorting: Research Technologyjessica hernandezPas encore d'évaluation
- CORMIC App NoteDocument4 pagesCORMIC App NoteRaja GopalPas encore d'évaluation
- Ajman 2019Document6 pagesAjman 2019Mohammad Asmat Ullah KhanPas encore d'évaluation
- Visualization of Cytoskeletal Elements by The Atomic Force MicroscopeDocument21 pagesVisualization of Cytoskeletal Elements by The Atomic Force MicroscopeBiraj KarmakarPas encore d'évaluation
- GE-INCell 3000Document15 pagesGE-INCell 3000api-19762689Pas encore d'évaluation
- BMC Neuroscience: Statistical Model of Natural Stimuli Predicts Edge-Like Pooling of Spatial Frequency Channels in V2Document12 pagesBMC Neuroscience: Statistical Model of Natural Stimuli Predicts Edge-Like Pooling of Spatial Frequency Channels in V2mano100% (1)
- Fractal Breathing of Voltage-Gated Ion ChannelDocument8 pagesFractal Breathing of Voltage-Gated Ion ChannelRenato MoraesPas encore d'évaluation
- Zhang-Zhao2019 Article FluorescenceMicroscopyImageClaDocument12 pagesZhang-Zhao2019 Article FluorescenceMicroscopyImageClaMahaPas encore d'évaluation
- BiophysicsDocument8 pagesBiophysicsPrabha SinghPas encore d'évaluation
- Bio Analytical TechniquesDocument26 pagesBio Analytical TechniquesanimalfacilityPas encore d'évaluation
- Term Paper On Transmission Electron MicroscopeDocument7 pagesTerm Paper On Transmission Electron Microscopeafdtveepo100% (1)
- Thesis On Comet AssayDocument4 pagesThesis On Comet Assaymichellebojorqueznorwalk100% (2)
- Accuracy of Direct Gradient Sensing by Single CellsDocument6 pagesAccuracy of Direct Gradient Sensing by Single CellsAntoan BekelePas encore d'évaluation
- Cellular StructureDocument12 pagesCellular StructureFekadu DagnawPas encore d'évaluation
- Survey Goals Segmentation: Watershed SegmentationDocument3 pagesSurvey Goals Segmentation: Watershed Segmentationjenisha SPas encore d'évaluation
- ATC: Lecture 4: Cell Separation Cell Quantification Cell CharacterizationDocument42 pagesATC: Lecture 4: Cell Separation Cell Quantification Cell Characterizationchan yi hanPas encore d'évaluation
- You Should Use Regression To Detect Cells: October 2015Document9 pagesYou Should Use Regression To Detect Cells: October 2015Vint PinePas encore d'évaluation
- Robust Off-And Online Separation of Intracellularly Recorded Up and Down Cortical StatesDocument11 pagesRobust Off-And Online Separation of Intracellularly Recorded Up and Down Cortical StatesdnloboPas encore d'évaluation
- Cell Cluster Graph For Prediction of Biochemical Recurrence in Prostate Cancer Patients From Tissue MicroarraysDocument11 pagesCell Cluster Graph For Prediction of Biochemical Recurrence in Prostate Cancer Patients From Tissue MicroarraysFrontiersPas encore d'évaluation
- Size Distributions of Intracellular Condensates Reflect Competition Between Coalescence and NucleationDocument22 pagesSize Distributions of Intracellular Condensates Reflect Competition Between Coalescence and NucleationonesimusPas encore d'évaluation
- Detection of Blockage in Coronary Artery in Coronary AngiographyDocument7 pagesDetection of Blockage in Coronary Artery in Coronary AngiographyDevi SriPas encore d'évaluation
- Retina Blood Vessel SegmentationDocument28 pagesRetina Blood Vessel SegmentationRevati WablePas encore d'évaluation
- Research Article Robust Blood Cell Image Segmentation Method Based On Neural Ordinary Differential EquationsDocument11 pagesResearch Article Robust Blood Cell Image Segmentation Method Based On Neural Ordinary Differential EquationsSamir GhoualiPas encore d'évaluation
- Cell Spreading Kinetics Guided by Interfacial StiffnessDocument11 pagesCell Spreading Kinetics Guided by Interfacial StiffnesssaawanPas encore d'évaluation
- IET Image Processing - 2023 - Zhou - Accurately 3D Neuron Localization Using 2D Conv LSTM Super Resolution SegmentationDocument14 pagesIET Image Processing - 2023 - Zhou - Accurately 3D Neuron Localization Using 2D Conv LSTM Super Resolution SegmentationAnu s sPas encore d'évaluation
- Konokhovaetal. 2013 High PrecisioncharacterizationofindividualE.cDocument8 pagesKonokhovaetal. 2013 High PrecisioncharacterizationofindividualE.cakpPas encore d'évaluation
- Automatic Vessel Extraction With Combined Bottom Hat and Match FilterDocument5 pagesAutomatic Vessel Extraction With Combined Bottom Hat and Match FilterBahareh Bozorg ChamiPas encore d'évaluation
- Pi Is 0006349518321933Document1 pagePi Is 0006349518321933ASP ValenciaPas encore d'évaluation
- Deep Learning-Based Cell Identification and Disease Diagnosis Using Spatio-Temporal Cellular Dynamics in Compact Digita PDFDocument18 pagesDeep Learning-Based Cell Identification and Disease Diagnosis Using Spatio-Temporal Cellular Dynamics in Compact Digita PDFJeffreyPas encore d'évaluation
- Self-organizing Neural Maps: The Retinotectal Map and Mechanisms of Neural Development: From Retina to TectumD'EverandSelf-organizing Neural Maps: The Retinotectal Map and Mechanisms of Neural Development: From Retina to TectumPas encore d'évaluation
- Educ 1301 Field Experience 1 ThielenDocument4 pagesEduc 1301 Field Experience 1 Thielenapi-610903961Pas encore d'évaluation
- Mental AspectDocument29 pagesMental AspectBenjii CarlosPas encore d'évaluation
- Grupo Stoncor Description - Stonhard Carboline Fibergrate PDFDocument22 pagesGrupo Stoncor Description - Stonhard Carboline Fibergrate PDFAndres OsorioPas encore d'évaluation
- EN6VC IIIa 6.2 - 2023 2024Document2 pagesEN6VC IIIa 6.2 - 2023 2024Ma. Feliza SaliganPas encore d'évaluation
- 5 Axis MachinesDocument33 pages5 Axis MachinesgsPas encore d'évaluation
- Audi A3 Injeção DiretaDocument109 pagesAudi A3 Injeção Diretawesley candido100% (1)
- IIT BOMBAY RESUME by SathyamoorthyDocument1 pageIIT BOMBAY RESUME by SathyamoorthySathyamoorthy VenkateshPas encore d'évaluation
- Theories of SelfDocument5 pagesTheories of SelfTd Devi AmmacayangPas encore d'évaluation
- LTE Interview QuestionsDocument10 pagesLTE Interview QuestionsMahesh Pratap100% (1)
- Arduino - Decision Making StatementsDocument20 pagesArduino - Decision Making StatementsJohn Clifford Ambaic JayomaPas encore d'évaluation
- Foundation ProblemsDocument71 pagesFoundation Problemsيقين يقين0% (1)
- Galvanized and Black Malleable Iron Pipe Fittings SpecificationsDocument24 pagesGalvanized and Black Malleable Iron Pipe Fittings SpecificationsdeniPas encore d'évaluation
- LADY BroshureDocument2 pagesLADY BroshureMcGabi GabrielPas encore d'évaluation
- Basf Masteremaco Application GuideDocument15 pagesBasf Masteremaco Application GuideSolomon AhimbisibwePas encore d'évaluation
- Trends in Structural Systems and Innovations for High-Rise Buildings over the Last DecadeDocument53 pagesTrends in Structural Systems and Innovations for High-Rise Buildings over the Last DecadeNarasimhaReddy PangaPas encore d'évaluation
- 07-02-2023 MS QLNDocument3 pages07-02-2023 MS QLNSummaaaaPas encore d'évaluation
- Optimization Module For Abaqus/CAE Based On Genetic AlgorithmDocument1 pageOptimization Module For Abaqus/CAE Based On Genetic AlgorithmSIMULIACorpPas encore d'évaluation
- Ethics in ResearchDocument21 pagesEthics in Researchmukku_raviPas encore d'évaluation
- DUPLICATE BILL TITLEDocument8 pagesDUPLICATE BILL TITLEMohammed AhmedPas encore d'évaluation
- Soft Computing Techniques Assignment1 PDFDocument14 pagesSoft Computing Techniques Assignment1 PDFshadan alamPas encore d'évaluation
- Writing Emails Part 1 Informal British English Teacher Ver2Document7 pagesWriting Emails Part 1 Informal British English Teacher Ver2Madalina MandiucPas encore d'évaluation
- Dayal Bagh: Spircon 2013: Book of AbstractsDocument202 pagesDayal Bagh: Spircon 2013: Book of AbstractsSant Mat100% (2)
- ZXJ 10 SPCDocument7 pagesZXJ 10 SPCMuhammad Yaseen100% (1)
- CitationCJ4-CB Checklists B v1.2Document4 pagesCitationCJ4-CB Checklists B v1.2Joao Vitor RojasPas encore d'évaluation
- The Scientific MethodDocument4 pagesThe Scientific MethodRob LovPas encore d'évaluation
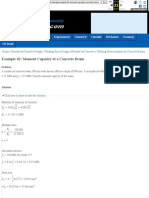
- Example 02 - Moment Capacity of A Concrete Beam - Reinforced Concrete Design ReviewDocument5 pagesExample 02 - Moment Capacity of A Concrete Beam - Reinforced Concrete Design ReviewJohn Michael GeneralPas encore d'évaluation