Académique Documents
Professionnel Documents
Culture Documents
Vertebrate Systems Summary
Transféré par
Hazel Grace BellenDescription originale:
Copyright
Formats disponibles
Partager ce document
Partager ou intégrer le document
Avez-vous trouvé ce document utile ?
Ce contenu est-il inapproprié ?
Signaler ce documentDroits d'auteur :
Formats disponibles
Vertebrate Systems Summary
Transféré par
Hazel Grace BellenDroits d'auteur :
Formats disponibles
BIO 342 Comparative Vertebrate Anatomy Lecture Notes 2 - Vertebrate Skeletal Systems
Source: http://www.auburn.edu/academic/classes/zy/0301/Topic3b/Topic3b.html
o
Bone:
after ostracoderms, fish continued to develop much bone in skin but that bone has become 'thinner' over time
inorganic components of bone comprise 60% of the dry weight (largely calcium hydroxy-appetite crystals) & provide the compressive strength of bone. The organic component is primarily collagen, which gives bone great tensile strength. provides support and movement via attachments for soft tissue and muscle, protects vital organs, is a major site for red marrow for production of blood cells, and plays a role in the metabolism of minerals such as calcium and phosphorus. There are two basic structural types of bone, compact and spongy. Compact bone forms the outer shell of all bones and also the shafts in long bones. Spongy bone is found at the expanded heads of long bones and fills most irregular bones. .
Endoskeleton o Somatic - axial & appendicular skeletons o Visceral - cartilage or bone associated with gills & skeletal elements (such as jaw cartilages) derived from them
Source: http://www.auburn.edu/academic/classes/zy/0301/Topic11/Topic11.html Dermal bone of fishes:
Bone formation begins with a blastema (any aggregation of embryonic mesenchymal cells which will differentiate into tissue such as muscle, cartilage, or bone). These cells then develop into either FIBROBLASTS or OSTEOBLASTS. Fibroblasts form collagen; osteoblasts form bone cells. Together, these form MEMBRANE BONE (bone deposited directed in a blastema).
Basic structure includes lamellar (compact) bone, spongy bone, dentin, &, often, a surface with a layer of enamel-like material Evolutionary 'trend' = large bony plates giving way to smaller, thinner bony scales o Ancient armor - not found on living fish o Ganoid scales - found only on Latimeria (coelocanth) & sturgeons o Placoid scales - elasmobranchs (diagram to the right; pulp cavity > dentin layer > enamel) o Ctenoid & Cycloid scales - modern bony fish
Intramembranous ossification is the process of membrane bone formation. This process give rise to:
bones of the lower jaw, skull, & pectoral girdle dentin & other bone that develops in the skin vertebrae in some vertebrates (teleosts, urodeles, & apodans)
Also check: http://courses.washington.edu/vertebra/453/photos/skin_photos/special_integume nt1.htm
Endochondral ossification is the process in which bone is deposited in preexisting cartilage, & such bone is called REPLACEMENT BONE.
1 = lamellar bone, 2 = spongy bone, 3 = dentin, 4 = enameloid, & 5 = fibrous plate (collagen) Source: http://www.uta.edu/biology/restricted/3452int.htm
Source: http://training.seer.cancer.gov/module_anatomy/unit3_3_bone_growth.html
Tetrapods - retain dermal elements in the skull, jaws, & pectoral girdle Somatic skeleton = axial skeleton (vertebral column, ribs, sternum, & skull) + appendicular skeleton Vertebral column: Vertebrae - consist of a centrum (or body), 1 or 2 arches, plus various processes
Skeletal elements:
Dermal skeleton o skin of most living vertebrates has no hard skeletal parts but dermal bone elements are usually present in the head region o early vertebrates (ostracoderms) had so much dermal bone they were called 'armored fishes'
Amphicelous o concave at both ends o most fish, a few salamanders (Necturus), & caecilians Opisthocoelous o convex in front & concave in back o most salamanders Procelous o concave in front & convex in back o anurans & present-day reptiles Acelous o flat-ended o mammals Heterocelous o saddle-shaped centrum at both ends o birds
Cartilaginous fishes o do not have typical fish vertebral columns o vertebrae include neural arches (cartilaginous dorsal plates) & dorsal intercalary plates are located between successive arches Teleosts
o o o
well-ossified amphicelous vertebrae notochord persists within each centrum (but constricted) neural arch associated with each centrum & hemal arches in tail (caudal) vertebrae
Chondrosteans (sturgeons & paddlefish) & modern lungfishes o incomplete centra o notochord is not constricted o cartilage deposited in notochord sheath provides structural support
Diplospondyly = 2 centra and 2 sets of arches per body segment; occurs in some fish (including sharks) Vertebral arches: Agnathans - only skeletal elements associated with the notochord are paired, lateral neural cartilages
Neural arch - on top of centrum Hemal arch (also called chevrons) - beneath centrum in caudal vertebrae of fish, salamanders, most reptiles, some birds, & many long-tailed mammals
Vertebral columns of tetrapods
Vertebral processes:
projections from arches & centra some give rigidity to the column, articulate with ribs, or serve as sites of muscle attachment
Cervical region o Amphibians - single cervical vertebra; allows little head movement o Reptiles - increased numbers of cervical vertebrae (usually 7) & increased flexibility of head o Birds - variable number of cervical vertebrae (as many as 25 in swans)
o o
Transverse processes - most common type of process; extend laterally from the base of a neural arch or centrum & separate the epaxial & hypaxial muscles Diapophyses & parapophyses - articulate with ribs Prezygapophyses (cranial zygapophyses) & postzygapophyses (caudal zygapophyses) - articulate with one another & limit flexion & torsion of the vertebral column
Mammals - usually 7 cervical vertebrae Reptiles, birds, & mammals - 1st two cervical vertebrae are modified & called the atlas & axis atlas - 1st cervical vertebra; ring-like (most of centrum gone); provides 'cradle' in which skull can 'rock' (as when nodding 'yes') axis - 2nd cervical vertebra Transverse foramen (#6 in above caudal view of a cervical vertebra) found in cervical vertebrae of birds & mammals provides canal for vertebral artery & vein
Vertebral columns:
Dorsal region
o o
Dorsals - name given to vertebrae between cervicals & sacrals when all articulate with similar ribs (e.g., fish, amphibians, & snakes) Crocodilians, lizards, birds, & mammals - ribs are confined to anterior region of trunk thoracic - vertebrae with ribs lumbar - vertebrae without ribs
The 'trend' in vertebra evolution has been for pleurocentra to increase in size (and, of course, for the hypocentrum to decrease in size). This trend is apparent in this diagram:
Cross-hatched areas = hypocentrum; black areas = pleurocentrum. Ribs - may be long or short, cartilaginous or bony; articulate medially with vertebrae & extend into the body wall
Sacrum & Synsacrum o sacral vertebrae - have short transverse processes that brace the pelvic girdle & hindlimbs against the vertebral column Amphibians - 1 sacral vertebra Living reptiles & most birds - 2 sacral vertebrae Most mammals - 3 to 5 sacral vertebrae o Sacrum - single bony complex consisting of fused sacral vertebrae; found when there is more than 1 sacral vertebra (see examples below):
A few teleosts - have 2 pair of ribs for each centrum of trunk (dorsal rib separates epaxial & hypaxial muscles)
Synsacrum found in birds produced by fusion of last thoracics, all lumbars, all sacrals, & first few caudals fused with pelvic girdle provides rigid support for bipedal locomotion
Caudal region o Primitive tetrapods - 50 or more caudal vertebrae o Present-day tetrapods number of caudal vertebrae is reduced arches & processes get progressively shorter (the last few caudals typically consist of just cylindrical centra as shown below) o Anurans - unique terminal segment called the urostyle (section of unsegmented vertebral column probably derived from separate caudals of early anurans) o Birds - last 4 or 5 caudal vertebrae fused to form pygostyle (see drawing above) o Apes & humans - last 3 to 5 caudal vertebrae fused to form coccygeal (or tail bone)
Most teleosts - ventral ribs only Sharks - dorsal ribs only Agnathans - no ribs Tetrapods - ribs usually articulate with vertebrae in moveable joints (see above drawing) o Early tetrapods - ribs articulated with every vertebra from the atlas to the end of the trunk o Later tetrapods - long ribs limited to thoracic region Thoracic ribs - most composed of a dorsal element (vertebral rib) & a ventral element (sternal rib) Sternal rib - may be ossified (birds) or remain cartilaginous (mammals); usually articulate with sternum (except 'floating ribs') Uncinate processes - found in birds; provides rib-cage with additional support
Sternum - strictly a tetrapod structure &, primarily, an amniote structure.
Amphibians - no sternum in early amphibians &, among present-day amphibians, only anurans have one Amniotes o sternum is a plate of cartilage & replacement bone o sternum articulates with the pectoral girdle anteriorly & with a variable number of ribs
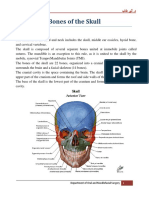
Useful link: BIO 342 Comparative Vertebrate Anatomy Lecture Notes - Skeletal System II (Skull) The Vertebrate Skull consists of: 1 - neurocranium (also called endocranium or primary braincase)
EVOLUTION OF VERTEBRAE
Unlike most tetrapods today, vertebral column of earliest tetrapods did not consist of 1 bone/body segment. Crossopterygian vertebrae consisted of an hypocentrum (a large, wedgeshaped piece) plus 2 pleurocentra (smaller, intersegmental pieces). This type of vertebra is called a rachitomous vertebra.
2 - dermatocranium (membrane bones) 3 - splanchnocranium (or visceral skeleton)
Neurocranial ossification centers: 1 - occipital centers
Neurocranium: 1 - protects the brain 2 - begins as cartilage that is partly or entirely replaced by bone (except in cartilaginous fishes)
Cartilaginous stage: neurocranium begins as pair of parachordal & prechordal cartilages below the brain parachordal cartilages expand & join; along with the notochord from the basal plate prechordal cartilages expand & join to form an ethmoid plate Cartilage also appears in the olfactory capsule (partially surrounding the olfactory epithelium) otic capsule (surrounds inner ear & also develops into sclera of the eyeball)
cartilage surrounding the foramen magnum may be replaced by as many as four bones: o basioccipital o exoccipital (2) o supraoccipital Mammals - all 4 occipital elements typically fuse to form a single occipital bone (pictured below) Tetrapods - neurocranium articulates with the 1st vertebra via 1 (reptiles and birds) or 2 (amphibians and mammals) occipital condyles (see human skull below)
2 - Sphenoid centers form:
basisphenoid bone (anterior to basioccipital) presphenoid bone side walls above basisphenoid & presphenoid form: o orbitosphenoid o pleurosphenoid o alisphenoid
Completion of floor, walls, & roof: Ethmoid plate - fuses with olfactory capsules Basal plate - fuses with otic capsules Further development of cartilaginous neurocranium = development of cartilaginous walls (sides of braincase) &, in cartilaginous fishes, a cartilaginous roof over the brain
3 - Ethmoid centers tend to remain cartilaginous & form
anterior to sphenoid cribiform plate of ethmoid & several conchae (or ethmoturbinal bones)
Cartilaginous fishes - retain a cartilaginous neurocranium (or chondrocranium) throughout life Bony fishes, lungfishes, & most ganoids - retain highly cartilaginous neurocranium that is covered by membrane bone Cyclostomes - the several cartilaginous components of the embryonic neurocranium remain in adults as more or less independent cartilages The ethmoid region is clearly visible within the bisected skull above. In most mammals, the nasal chamber is large & filled with ridges from the ethmoid bones called the turbinals or ethmoturbinals. These bones are covered with olfactory epithelium in life and serve to increase the surface area for olfaction (i.e., a more acute sense of smell). Another ethmoid bone, the cribiform plate, separates the nasal chamber from the brain cavity within the skull. 4 - Otic centers - the cartilaginous otic capsule is replaced in lower vertebrates by several bones:
Source: http://gwis2.circ.gwu.edu/~atkins/newwebpages/Skeletal/Skeletable.html Other bony vertebrates - embryonic cartilaginous neurocranium is largely replaced by replacement bone (the process of endochondral ossification occurs almost simultaneously at several ossification centers)
prootic opisthotic
epiotic One or more of these may unite with adjacent replacement or membrane bones: o Frogs & most reptiles - opisthotics fuse with exoccipitals o Birds & mammals - prootic, opisthotic, & epiotic unite to form a single petrosal bone; the petrosal, in turn, sometimes fuses with the squamosal to form the temporal bone
Sharks - cartilaginous bony vertebrates - membrane bones form Birds, mammals, & some reptiles - a secondary ('false') palate develops creating a horizontal partition that separates the oral cavity into nasal & oral passages. The secondary palate is formed from processes of the premaxillae, maxillae, and palatines.
Opercular bones
DERMATOCRANIUM - lies superficial to neurocranium & forms: 1 - bones that form the roof of the brain & contribute to the lateral walls of the skull 2 - bones of the upper jaw 3 - bones of the palate(s) 4 - opercular bones
Operculum = fold of the hyoid arch that extends back over the gill slits in holocephalans & bony fishes Tetrapods - no vestiges of opercular bones remain
BIO 342 Comparative Vertebrate Anatomy Lecture Notes 4 - Skeletal System III VISCERAL SKELETON (or SPLANCHNOCRANIUM)
skeleton of the pharyngeal arches o Fishes - skeleton of the jaws & gill arches o Tetrapods - skeleton modified for new functions
Basic pattern of "roofing bones":
crossopterygians - a series of paired & unpaired bones along mid-dorsal line of skull (below left) labyrinthodonts - unpaired bones lost & a series of paired bones resulted (nasals, frontals, parietals, & dermoccipitals) (below right) Fontanels = 'soft spots' o occur when neurocranium is incomplete dorsally (e.g., teleosts & tetrapods) o can be felt in head until the membranes under the skin have ossified
Fish visceral skeleton - consists of 7 sets of paired cartilages in the 7 visceral arches & a series of mid-ventral cartilages (basihyal & basibranchials) in the pharyngeal floor
Bony fishes
Bones of the upper jaw
Pterygoquadrate (palatoquadrate) cartilage - 1st upper jaw that vertebrate embryos develop o Cartilaginous fishes - palatoquadrate is the only upper jaw that develops o Bony vertebrates - the palatoquadrate becomes covered with dermal bones (premaxillae & maxillae) that make up the adult upper jaw
visceral skeleton resembles that of sharks except that bone is added caudal ends of the cartilaginous pterygoquadrate undergo endochondral ossification & become the quadrate bones. The remainder becomes the palatine & pterygoid bones. The posterior tip of Meckel's cartilage becomes an articular bone. (See http://www.usm.maine.edu/bio/courses/bio205/bio205_08_skull_2.html)
Feeding movements in many bony fishes -> cranial kinesis (see Figure 9.23, p.351 of text) Cranial kinesis:
Palatal bones - the floor on which the brain rests is at the same time the roof of the oral cavity in fishes & amphibians (primary palate)
movement between the upper jaw and braincase advantages:
provides a way to change the size and configuration of the mouth rapidly o optimize biting and rapid feeding. disadvantages: lose force, difficult to optimize apposition of occlusive surfaces.
o o
Reptiles - largely ensheathed by dermal bones (as in the above turtle) Birds & mammals - few or no remnants in adult lower jaw (&, in mammals, the articular, formed by ossification of the tip of Meckel's cartilage, projects into the middle ear cavity & becomes the malleus)
See also: Shark Jaw Movement Source: http://home.houston.rr.com/vnotes/Bones/Meckel.html Cyclostomes
visceral skeleton unlike that of jawed fishes Hagfishes - no identifiable pterygoquadrate or Meckel's cartilage
Jaw suspension of fishes
Arch II = Hyomandibular cartilage: o Sharks - interposed between quadrate region & otic capsule o Tetrapods - no longer articulates with quadrate & ossifies to become part of the stapes (columella) Arches III ---> V become part of hyoid apparatus Arches VI & VII - not present in tetrapods
The jaw-hyoid complex of fishes requires bracing against some support to function effectively, and the nearest one is the neurocranium (endocranium). Types of suspensions: o autostyly (below left) - hyomandibula play no role in bracing the jaws (lungfish & tetrapods) o amphistyly (below middle) - jaws & hyomandibula both braced directly against the braincase (extinct sharks) o hyostyly (below right) - mandibular cartilage is braced against the otic capsule; jaws braced against hyomandibula (sharks & present-day bony fishes) Source: http://www.uta.edu/biology/restricted/3452hske.htm
Source: http://www.liberty.edu/academics/registrar/courses00/biol418pa.htm Hyoid apparatus of tetrapods
consists of a body & 2 or 3 horns (cornua) anchors tongue, provides attachment for some extrinsic muscles of larynx, & is site of attachment of muscles that aid in swallowing
Lower jaw
TETRAPODS - With life on land (& pulmonary respiration), the visceral skeleton underwent substantial modification. Some structures were lost & others remained to perform new functions.
may have originated as part of a visceral arch, as in sharks (mandibular cartilage) in bony vertebrates, mandibular cartilage is reinforced & largely replaced by a series of dermal bones
Pterygoquadrate (palatoquadrate) cartilage = embryonic upper jaw cartilage o Amphibians, reptiles, & birds - posterior end undergoes endochondral ossification & becomes the quadrate (which articulates with the articular bone of the lower jaw)
Skeletal BIO 342 Comparative Vertebrate Anatomy Lecture Notes 5 - Skeletal System IV Appendicular Skeleton Appendicular skeleton
Source: http://www-users.york.ac.uk/~wjh101/hedbone/Turtle/turtle.htm Mammals - dentary (lower jaw) articulates with the squamosal of skull (quadrate separates from the rest of the palatoquadrate & becomes the incus of the middle ear) Meckel's cartilage
consists of pectoral & pelvic girdles plus skeleton of fins & limbs Some vertebrates have no appendicular skeleton (e.g., agnathans, apodans, snakes, & some lizards) & in others it is much reduced.
Pectoral girdles: 1 - brace for anterior appendages 2 - consist of membrane & replacement bones (in bony vertebrates) 3 - Early fishes had 3 replacement bones (coracoid, scapula, & suprascapula) and a series of dermal bones (clavicle, cleithrum, supracleithrum, and post-temporal) 4 - Later bony fishes (ganoid fish) - tendency for reduction in number and size of replacement bones 5 - Tetrapods - tendency for reduction in number of dermal bones
Fishes - pelvic girdle consists of 2 cartilaginous or bony plates (ischiopubic plates) that articulate with the pelvic fins
Source: http://www.uta.edu/biology/restricted/3452gird.htm
Tetrapods - pair of cartilaginous plates form in embryos & each ossifies at 2 centers to form: pubis & ischium. An additional blastema gives rise to the ilium.
Frogs & toads
Source: http://www.uta.edu/biology/restricted/3452gird.htm Bony fishes - pectoral girdles of living bony fishes have reduced coracoid & scapula (replacement bone) but large cleithrum & supracleithrum (dermal bone). A posttemporal bone (dermal) connects the supracleithrum to the skull.
ilia elongated & extend from sacral vertebra to urostyle joint between ilium & sacral vertebra (sacroiliac) is freely moveable (& moves when a frog or toad jumps)
Source: http://www.uta.edu/biology/restricted/3452gird.htm Cartilaginous fishes - no dermal bone
Birds:
Source: http://www.uta.edu/biology/restricted/3452gird.htm Tetrapods - early ones had pectoral girdle similar to those of early bony fishes, but lost posttemporal & acquired interclavicle (which still occurs in several amniotes, e.g., alligator, birds, & monotremes) Clavicle & coracoid - one or both typically brace scapula against sternum (as in birds; below) Scapula - present in all tetrapods with even vestiges of anterior limbs, e.g., turtles & birds & mammals
ilium & ischium expanded to accommodate musculature needed for bipedal locomotion girdle is braced against lumbar & sacral vertebrae pubic bones are typically reduced (long but thin); the limited pubic symphysis provides a larger outlet for eggs
Mammals - ilium, ischium, and pubis unite to form the innominate bone (the 2 innominates = pelvic girdle) FINS
Pelvic girdles
brace posterior paired appendages no dermal components (unlike pectoral girdle)
All jawed fishes (except eels) have pectoral & pelvic fins Fins are used primarily for steering ('rudders') Types of fins: o lobed fins - found in sarcopterygians o fin fold fins (see diagram below) found in cartilaginous fish consist of 1 to 5 basal cartilages plus several rows of radials o ray fin - tendency to lose proximal components of fin skeleton (see diagram below)
Limbs
Starting with amphibians, vertebrates typically have 4 limbs. However, some have lost one or both pairs &, in others, one pair is modified as arms, wings, or paddles typically have 5 segments: o Anterior limb brachium (upper arm) - consists of humerus antebrachium (forearm) - consists of radius & ulna carpus (wrist) - consists of carpals metacarpus (palm) - consists of metacarpals digits - consist of phalanges o Posterior limb femur (thigh) - consists of femur crus (shank) - consists of tibia & fibula tarsus (ankle) - consists of tarsals metatarsus (instep) - consists of metatarsals digits - consist of phalanges
distal row = 5 distal carplas numbered 1 through 5 (starting on thumb, or radial, side) Palm - metacarpals Digits each consists of a series of phalanges general formula' (starting at thumb) = 2,3,4,5,3
Adaptive modifications of the manus 1 - Flight
Birds - loss of digits & bones plus fusion of some bones Bats - 5 digits; elongated metacarpals (II through V) & phalanges support the patagium Pterosaurs - 4th digit elongated to support patagium
Source: http://www.uta.edu/biology/restricted/3452gird.htm Some vertebrates lack both pairs of limbs
2 - Swimming - increase in number, & size, of phalanges
3 - Terrestrial locomotion (walking & running):
caecilians (apodans) most snakes snake-like lizards
Some vertebrates have forelimbs only:
manatees & dugongs dolphins (see diagram below) cetaceans (vestigial elements may be embedded in body wall) sirens (salamander)
Plantigrade flat-footed all bones of manus and/or pes on the ground amphibians, most reptiles (see alligator photo above), & some mammals (insectivores, monkeys, apes, humans, & bears) Digitigrade 1st digit is reduced or lost manus & pes are elevated rabbits, rodents, & many carnivores Unguligrade reduced number of digits walk on tips of remaining digits claws become hooves
Early tetrapods - limbs were short & first segment extended straight out from the body . This posture persists among lower tetrapods (e.g, see the alligator below), but, in birds & mammals, there has been a rotation of the appendages so that the long axis of the humerus & femur more nearly parallels the vertebral column.
Arm & forearm o Upper arm = humerus o Forearm = radius & ulna o Manus (or hand) Wrist - 3 rows of carpal bones: proximal row = radiale, ulnare, intermedium, & pisiform middle row = 3 central carpals (centralia)
As the fastest North American mammal, pronghorn antelope (unguligrades) can reach speeds of 60 miles per hour. At high speed they cover the ground in strides of 14 to 24 feet, and are known to run for long distances at speeds of 30 to 40 miles per hour.
4 - Grasping
E. Ichthyostega, F. Acanthostega
Here we have a possible explanation for opposable thumb the formation of a new morphological saddle joint at base of thumb where it meets palm feature - limbs with digits - from the paired fins of sarcopterygian (lobe thumb at wider angle from index finger finned) fishes. Some of these fish strong thumb muscles (notably Eusthenopteron) have bones in their paired fins that are very similar to Posterior limbs - bones are comparable to those of forelimbs except that a patellabones of tetrapod limbs. the (kneecap') develops in birds & mammals Specifically, they have a single bone (similar to the humerus or femur) followed by paired bones (similar to the Origin of fins: radius and ulna or fibula and tibia of tetrapods). Scientists think Eusthenopterons used their limbs to 1 - Fin fold hypothesis - fins derived from continuous fold of body wall walk on the sea (or lake or river) bed. What Eusthenopteron lacks are digits having fin rays instead (although Ammoecetes Sauripterus is a closely related fish that does have 8 digits just like the earliest amphibians - see the illustration). 2 - Gill arch hypothesis - fins derived from the last 2 gill arches (very unlikely) However, the fossil record supplies us with examples of tetrapods that are 3 - Fin spine hypothesis - fins derived from tissue attached to spines (that may quite similar to Eusthenopteron have evolved to provide protection from predators) Acanthostega and Icthyostega.
Acanthodiian
BIO 342 Comparative Vertebrate Anatomy Lecture Notes 6 - Muscular System Vertebrate muscles:
Origin of limbs - derived from paired fins of ancient fishes: Used by permission of John W. Kimble
striated vs. smooth voluntary vs. involuntary skeletal vs. non-skeletal
The first tetrapods, Labyrinthodont amphibians (right), probably evolved from a Crossopterygian ancestor (left). When the fresh water pools in which these fish lived became stagnant, they may have crawled up the bank to breath air using primitive lungs. As the lobed fins of these fish evolved into stronger limbs, the first tetrapods appeared.
Skeletal muscle (left) & Smooth muscle (right) Skeletal muscle = muscles attached to the skeleton that are striated & voluntary Non-skeletal muscle = muscles not attached to the skeleton; most are smooth & involuntary
arison of paired anterior fins of nned fishes (A-D) and limbs of etrapods (E, F)
Vertebrate Muscles: 1 - Skeletal, striated, voluntary muscles
ropterygion, B. Sauripterus, derichthys, D. Eusthenopteron,
axial
o o o
body wall & tail hypobranchial & tongue extrinsic eyeball muscles appendicular branchiomeric (homologous to the branchial/ pharyngeal muscles from fishes to mammals, striated muscles, innervated by cranial nerves) integumentary
Human muscles were named several hundred years ago, & many of these names are still used. Based on similarities of origins & insertions, these names were subsequently used for the, apparently, corresponding muscles of other vertebrates. However, origins & insertions are not reliable criteria for determining homology because natural selection has sometimes favored 'shifts' in muscle position. More reliable criteria for determining homologies are:
o o
embryonic origin nerve supply
2 - Non-skeletal, smooth, chiefly involuntary muscles Source: http://www.ucalgary.ca/UofC/eduweb/virtualembryo/why_fish.html
o o o
muscles of tubes, vessels, & hollow organs intrinsic eyeball muscles erectors of feathers & hair
Axial Muscles:
3 - Cardiac muscle 4 - Electric organs Skeletal muscles have muscular & tendinous portions:
Muscle - consists of skeletal muscle cells (which, in turn, consist of myofibrils and myofilaments) Tendons - extensions of a muscle's tough connective tissue sheath (fascia & epimysium) that anchor a muscle to its origin & insertion
include the skeletal muscles of the trunk & tail extend forward beneath the pharynx as hypobranchial muscles & muscles of the tongue are present in orbits as extrinsic eyeball muscles (check slide 27 in this powerpoint presentation) are metameric (most evident in fish and aquatic amphibians where the axial muscles are used in locomotion; in other tetrapods, metamerism is obscured due to presence of paired appendages responsible for locomotion on land) are segmental because of their embryonic origin; arise from segmental mesodermal somites
o o
Origin = site of attachment that is relatively fixed Insertion = site of attachment that is normally displaced by contraction of the muscle
Source: http://www.uta.edu/biology/restricted/3452mus.htm
Used with permission of John W. Kimball Names of skeletal muscles are based on:
Axial musculature of an aquatic salamander, Necturus maculosus. The layers of lateral hypaxial musculature are exposed from superficial to deep in the cranial to caudal direction. The number of external oblique layers varies between one and two in this species (the figured specimen exhibits two). Abbreviations: oes, M. obliquus externus superficialis; oep, M. obliquus externus profundus; oi, M. obliquus internus; ta, M. transversus abdominis (Brainerd and Simons 2000). Trunk & tail muscles of fish: Axial musculature consists of a series of segments (myomeres) separated by myosepta
direction of fibers (e.g., oblique) location or position (e.g., superficial) number of divisions (e.g., triceps) shape (e.g., deltoid) origin and/or insertion (e.g., iliocostalis) action (e.g., levator scapulae) size (e.g., major) or some combination of these
o o
Homologies:
Myosepta serve as origins & insertions for segmented muscles Myomeres are divided into dorsal & ventral masses by a horizontal septum that extends between the transverse processes of the vertebrae: Epaxials = above the septum Hypaxials = below the septum
Middorsal & midventral septa separate the myomeres of the 2 sides of the body. The midventral septum is called the LINEA ALBA.
lies close to neural arches connects spinous processes or transverse processes with those several vertebrae anteriorly
Trunk & tail muscles of tetrapods Shortest bundles - intervertebrals
Tetrapods, like fish, have epaxial & hypaxial masses, & these retain some evidence of metamerism even in the highest tetrapods. Modifications:
Hypaxials of tetrapods: 1 - Muscles of lateral body wall:
remain segmented connect processes (spinous, transverse, & zygapophyses) of adjacent vertebrae
1 - epaxials are elongated bundles that extend through many body segments & that are located below the expanded appendicular muscles required to operate the limbs 2 - hypaxials of the abdomen have no myosepta & form broad sheets of muscle 3 - hypaxials are oriented into oblique, rectus, & transverse bundles Epaxials of tetrapods:
oblique (external & internal), transverse, & rectus muscles
2 - Muscles that form longitudinal bands in roof of body cavity (subvertebral muscles) Oblique & transverse muscles:
lie along vertebral column dorsal to transverse processes & lateral to neural arches extend from base of the skull to tip of the tail o Urodeles & some lizards - epaxials are obviously metameric & are referred to as the dorsalis trunci (see salamander above) o Higher tetrapods - superficial epaxial bundles form long muscles that extend over many body segments; deep bundles are still segmented
Longest bundles: 1- longissimus group
Early amphibians & reptiles o ribs developed in myosepta along entire length of the trunk o urodeles still have myosepta the length of the trunk, but ribs no longer form in all of them Modern amniotes o myosepta & ribs are restricted to the thorax (so abdominal muscles are not obviously segmented) o hypaxials form 3 layers: external oblique, internal oblique, & transverse (in the thorax region: external & internal intercostals, which play an important role in respiration, & transverse muscle)
lies on transverse processes of vertebrae; includes the longest epaxial bundles subdivisions include: longissimus dorsi longissimus cervicis longissimus capitis
1 - External intercostal muscles, 2 - Internal intercostal muscles, 3 - Ribs, 4 Intercartilaginous muscles, 5 - Sternum, 6 - Subcostal muscles, & 7 - Vertebral column Source: http://www.sci.port.ac.uk/rad/anatomy/07/006.htm Rectus muscles:
2 - iliocostalis group
3 - spinalis group
lateral to longissimus & spinalis arises on ilium & inserts on dorsal ends of ribs or uncinate processes
weakly developed in most fish; 'stronger' in tetrapods support ventral body wall & aid in arching the back in mammals - rectus abdominis (typically extends from the anterior end of the sternum to the pelvic girdle)
Subvertebral muscles:
underneath & against transverse processes of vertebrae includes the psoas & iliacus in the lumbar region & the longus colli in the neck less developed in the thorax & none in the tail
the tongue of amniotes is a 'sac' anchored to hyoid skeleton & filled with hypobranchial muscle
FUNCTION OF EPAXIALS OF TETRAPODS: 1 - short epaxials perform same function as in fish (side-to-side movements of vertebral column) 2 - short & long bundles arch & support the vertebral column 3 - most anterior bundles = attach to & move the skull
The neck muscles ending in "hyoid" are associated with the hyoid apparatus, whereas those beginning or ending with "thyro" are attached to the larynx. These muscles are hypobranchia and function in movement of the hyoid apparatus, larynx and/or floor of the mouth. Appendicular muscles - move fins or limbs
Extrinsic - originate on axial skeleton or fascia or trunk & insert on girdles or limbs Intrinsic - originate on girdle or proximal skeletal elements of appendage & insert on more distal elements
FUNCTION OF HYPAXIALS OF TETRAPODS: 1 - Aquatic urodeles = used chiefly for swimming Fish - appendicular muscles serve mostly as stabilizers; intrinsic muscles are limited in number & undifferentiated Tetrapods - appendicular muscles are much more complicated than in fish Diagram showing the onset and duration of lateral hypaxial muscle EMG activity relative to maximum body bending in two Ambystoma tigrinum during swimming. Values are means S.D. for alpha-burst (filled bars) and beta-burst (open bars) onset and duration times. OES, m. obliquus externus superficialis; OEP, m. obliquus externus profundus; OI, m. obliquus internus; TA, m. transversus abdominis. *Denotes the side of the body on which the electrodes were implanted (Bennett et al. 2001).
o o
greater leverage required for locomotion on land jointed appendages (as opposed to fins) require complex muscles
Extrinsic appendicular musculature
2 - Terrestrial urodeles = assist in locomotion 3 - Other tetrapods = reduced in volume compared to fish (because of shift in mode of locomotion); now support contents of abdomen, assist in respiration (especially intercostal muscles), & assist epaxials in bending vertebral column (rectus muscles)
Dorsal group of the forelimbs, e.g., trapezius and latissimus dorsi, arise on:
1 - fascia of trunk in lower tetrapods 2 - skull, vertebral column, & ribs to a point well behind the scapula in higher tetrapods & converge on the girdle & limb
Hypobranchial & tongue muscles:
Ventral group, e.g., pectoralis, arises on sternum & coracoid, & converge on limb
Fish hypobranchials extend forward from pectoral girdle & insert on mandible, hyoid, & gill cartilages o hypobranchials strengthen floor of pharynx & assist branchiomeric muscles in elevating floor of mouth, lowering jaw, & extending gill pouches Tetrapods o hypobranchials stabilize & move hyoid apparatus & larynx
RESULT = pectoral girdle & limb are joined to trunk by extrinsic appendicular muscles as illustrated in this diagram:
The 'muscular sling' of tetrapods. Appendicular muscles of the forelimbs suspend the anterior body of tetrapods from the shoulders. Some of these muscles are axial
muscles (rhomboideus & serratus ventralis), some are branchial muscles (trapezius), & some arise from the forelimb musculature itself (pectoralis). The pelvic girdle requires no such muscular anchoring because it is attached directly to the vertebral column. As a result, the volume of extrinsic muscle is relatively small in posterior limbs. Extrinsic appendicular muscles: 1 - most develop from hypaxial blastemas in the body wall 2 - referred to as secondary appendicular muscles because it was not their original function to operate appendages 3 - chief extrinsic muscles of forelimbs of tetrapods include: scapular deltoid, latissimus dorsi, rhomboideus, serratus ventralis, & pectorals Intrinsic appendicular muscles: 1 - form from blastemas within the limb bud 2 - called primary appendicular muscles
o o
Squalus & other fish - operate the jaws (adductor mandibulae & intermandibularis) Tetrapods muscles of 1st arch still operate jaws adductors of mandible: masseter & temporalis (see diagram below) pterygoid digastric
Muscles of the Hyoid Arch:
o o o
move hyoid arch aid in hearing (stapedial muscle) assist in moving lower jaw (e.g., digastric)
Muscles of 3rd & successive arches:
o o o
Appendicular muscles: Amphibians - much more complex than in fish Reptiles - more numerous & diverse than in amphibians; better support of body & increased mobility of distal segments of the limbs Birds - intrinsic musculature is reduced; pectoralis (downstroke muscle) & supracoracoideus (upstroke muscle) are enlarged Mammals - similar to reptiles but more diverse Branchiomeric muscles: 1 - associated with the pharyngeal arches 2 - series of skeletal & smooth muscles 3 - adductors, constrictors, & levators operate jaws plus successive gill arches Electric organs: Muscles of the Mandibular Arch:
Squalus - constrictors above & below gill chambers plus levators (including the cucullaris) that compress & expand the gill pouches Bony fish - muscles reduced; operculum plays important role in respiration Tetrapods - muscles further reduced; primary muscles include: stylopharyngeus (Arch III) - used for swallowing intrinsic muscles of the larynx or 'voicebox' (remaining arches) cucullaris - gives rise to trapezius, cleidomastoid, & sternocleidomastoid muscles of amniotes
Integumentary muscles: Extrinsic integumentary muscles (e.g., platysma)
o o o
originate (usually) on the skeleton & insert on the underside of the dermis striated move skin of amniotes
Intrinsic integumentary muscles (arrector pili muscles)
o o o
entirely within the dermis found in birds & mammals mostly smooth muscles
1 - consist of a number of electric discs (up to 20,000) piled in either vertical or horizontal columns
2 - each disc (electroplax) is a large coin-shaped cell 3 - evolved several times in a variety of fish (good example of convergent evolution)
readily absorbed (e.g., hummingbirds) vs. requiring extensive enzymatic activity (e.g., carnivores) constant food supply (e.g., herbivores) vs. scattered supply (e.g., carnivores)
The embryonic digestive tract of vertebrates consists of 3 regions: Several species of fish have evolved an electric organ in their tail that produces a continous electric signal that propogates through the water. These fish have specialized receptors on their skin surface that can "feel" electricity. Objects in their environment, such as rocks, plants and pirhanas disturb the flow of their electric signal through the water. This disturbance will affect how strong the electric field is on a patch of skin near the object. In other words, a non-conductive object such as a rock will cast an electric "shadow" on the skin. The skin receptors near the object sense these disturbances and then increase or decrease their signal rate (depending wether the local electric field increased or decreased in intensity). These signals are then sent up to specialized regions of the brain that collate all the information and compute a coherent "picture" of the fish's environment (see http://soma.npa.uiuc.edu/labs/nelson/electrolocation.html). Functions of electric organs: 1 - defense 2 - communication 3 - locating prey (electrolocation) BIO 342 Comparative Vertebrate Anatomy Lecture Notes 7 - Digestive System Digestive tract - tube from mouth to vent or anus that functions in: 1 - midgut - contains yolk or attached yolk sac 2 - foregut - oral cavity, pharynx, esophagus, stomach, & small
intestine
3 - hindgut - large intestine & cloaca Mouth & oral cavity. The oral cavity begins at the mouth & ends at the pharynx. Fish have a very short oral cavity, while tetrapods typically have longer oral cavities. The mammalian mouth is specialized to serve as a suckling and masticatory organ (with muscular cheeks).
Palate = roof of the oral cavity o primary palate - internal nares lead into the oral cavity anteriorly o secondary palate - nasal passages are located above the secondary palate and open at the end of the oral cavity
Teeth are derivations of dermal armor.
Placoid scales - show gradual transition to teeth at the edge of the jaw Composition of teeth - primarily dentin surrounded by enamel Vary among vertebrates in number, distribution in the oral cavity, degree of permanence, mode of attachment, & shape
Toothless vertebrates are found in every class of vertebrates and include agnathans, sturgeons, some toads, turtles, birds, & baleen whales.
ingestion digestion absorption egestion
A right whale swims at or near the surface of the water with its mouth open. Water and food enter through a gap in the front baleen plates, and food is caught in the matted baleen fringes inside. Toothed vertebrates:
Major subdivisions include the oral cavity, pharynx, esophagus, stomach, small & large intestines, and cloaca. Accessory organs include the tongue, teeth, oral glands, pancreas, liver, & gall bladder.
Differences in the anatomy of vertebrate digestive tracts is often correlated with the nature & abundance of food:
Fish - teeth are numerous & widely distributed in the oral cavity & pharynx Early tetrapods - teeth widely distributed on the palate; most amphibians & some reptiles still have teeth on the vomer, palatine, & pterygoid bones Crocodilians, toothed birds, & mammals - teeth are limited to the jaws
TEETH: 1 - have tended toward reduced numbers & distribution 2 - most vertebrates (through reptiles) have succession of teeth 3 - most vertebrates (except mammals) replace teeth in waves (back to front; every other tooth) 4 - mammals generally develop 2 sets of teeth: milk (deciduous) teeth & permanent teeth
Using a keen sense of smell, anteaters are able to effectively track down ant nests on the forest floor. Once a nest is found, the mammal usually rips it open with its sharp foreclaws to expose its delectable contents. The anteater then proceeds to catch and eat the ants by repetitively flicking its long sticky tongue in and out of the nest. The giant anteater's unique tongue can measure as long as two feet (60 cm). Functions of vertebrate tongues:
Morphological variation in teeth:
vertebrates other than mammals - all teeth are shaped alike (homodont dentition) mammals - teeth exhibit morphological variation: incisors, canines, premolars, & molars (heterodont dentition) o incisors = cutting o canines = piercing & tearing o premolars & molars = macerating
capturing & gathering food (see woodpecker tongue below) taste manipulate fluids & solids in oral cavity swallowing thermoregulation grooming human speech
Oral glands - secrete a variety of substances including:
saliva Lubrication and binding: the mucus in saliva is extremely effective in binding masticated food into a slippery bolus that (usually) slides easily through the esophagus without inflicting damage to the mucosa. Saliva also coats the oral cavity and esophagus, and food basically never directly touches the epithelial cells of those tissues. o Solubilizes dry food: in order to be tasted (by taste buds), the molecules in food must be solubilized. o Oral hygiene: The oral cavity is almost constantly flushed with saliva, which floats away food debris and keeps the mouth relatively clean. Saliva also contains lysozyme, an enzyme that lyses many bacteria and prevents overgrowth of oral microbial populations. o Initiates starch digestion: in most species, amylase is present in saliva and begins to digest dietary starch into maltose. Amylase does not occur in the saliva of carnivores. o Provides alkaline buffering and fluid: this is of great importance in ruminants, which have non-secretory forestomachs. o Evaporative cooling: clearly of importance in dogs, which have very poorly developed sweat glands - look at a dog panting after a long run and this function will be clear. poison (lizards, snakes, and mammals) anticoagulant (vampire bats; video)
Tongue:
Gnathostome fish & primitive amphibians - tongue is a simple crescentshaped elevation in the floor of the oral cavity caused by the underlying hyoid skeleton & is called the primary tongue Most amphibians - primary tongue (or hypobranchial eminence) + glandular field (or tuberculum impar) ('stuffed' with hypobranchial musculature) Reptiles & mammals - primary tongue + glandular field (or tuberculum impar) + lateral lingual swellings (more hypobranchial muscle) Birds - lateral lingual swellings are suppressed & intrinsic muscle is usually lacking
Tongue mobility:
Turtles, crocodilians, some birds, & whales - tongue is largely immobilized in the floor of the oral cavity & cannot be extended Snakes, insectivorous lizards & amphibians, & some birds - tongue sometimes long and may move in and out of the oral cavity (seehttp://www.autodax.net/feedingmovieindex.html) Mammals - tongue is attached to the floor of the oral cavity (via the frenulum) but can still be extended out of the oral cavity
Pharynx - part of digestive tract exhibiting pharyngeal pouches (at least in the embryo) that may give rise to slits
Fish - pharynx is respiratory organ Tetrapods: o pharynx is the part of the foregut preceeding the esophagus & includes: glottis (slit leading into the larynx) openings of auditory (eustachian) tubes opening into esophagus
for absorption of water, minerals and nitrogen. Abomasum
The abomasum is the true stomach and the only site on the digestive tr produces gastric juices (HCl and the enzymes, pepsin and rennin). Ing remains here for 1 to 2 hours. The intestine is located between the stomach & the cloaca or anus & is an important site for digestion & absorption. Vertebrate intestines are differentiated to varying degrees into small & large intestines.
Mammals - an epiglottis is positioned over the glottis so that, when a mammal swallows, the larynx is drawn forward against the epiglottis & the epiglottis blocks the glottis (which prevents food or liquids from entering the trachea)
Source: http://www.stroke.cwc.net/niweb/faq.htm Esophagus:
a distensible muscular tube connecting the pharynx & the stomach may have diverticulum called the crop (see diagram of pigeon below)
Fishes - relatively straight & short intestine in cartilaginous fishes & in primitive bony fishes (lungfish & sturgeon). However, the intestine of cartilaginous fishes has a spiral valve.
Stomach = muscular chamber(s) at end of esophagus
serves as storage & macerating site for ingested solids & secretes digestive enzymes Vertebrate stomachs: o Cyclostomes - weakly developed; similar to esophagus o Fish, amphibians, & reptiles - increasing specialization (more differentiated from the esophagus) o Birds - proventriculus (glandular stomach) and ventriculus (muscular stomach, or gizzard)
Amphibians - intestines differentiated into coiled small intestine and short, straight large intestine Reptiles & Birds - coiled small intestines & a relatively short large intestine (that empties into the cloaca) Mammals - small intestine long & coiled and differentiated into duodenum, jejunum, & ileum. The large intestine is often relatively long (but not as long as the small intestine). A cecum is often present at the junction of the small & large intestines in herbivores.
Mammals - well-developed stomach; ruminants have multichambered stomachs: Reticulo-rumen (reticulum and rumen)
://www.uta.edu/biology/restricted/3452dig.htm
Source: http://www.uta.edu/biology/restricted/3452dig.htm
Reticulum and rumen are often discussed together since each compartment is Accessory organs - Liver, gall bladder, & pancreas separated by a low partition. Eighty percent of the capacity of the stomach is related to the reticulo-rumen. The contents of the reticulum and rumen intermix freely. The rumen is the main fermentation vat where billions of microorganismsLiver & gall bladder attack and break down the relatively indigestible feed components of the o liver produces bile which is stored in the gall bladder ruminant's diet. (cyclostomes, most birds, and some mammals, including cervids, have no gall bladder) o bile aids in digestion by emulsifying fats (breaking fats down into Omasum tiny particles that permits more efficient digestion by enzymes) After fermentation in the reticulum and rumen, food passes to the omasum. The Pancreas - secretes pancreatic juice (bicarbonate solution to neutralize omasum acts as a filter pump to sort liquid and fine food particles. Coarse fibre acids coming from the stomach plus enzymes to help digest carbohydrates, particles are not allowed to enter the omasum. Also, the omasum may be the site fats, and proteins) into the intestine
Ceca - blind diverticula that serve to increase the surface area of the vertebrate digestive tract
Fishes - pyloric & duodenal ceca are common in teleosts; these are primary areas for digestion and absorption (not fermentation chambers) Tetrapods - ceca are present in some herbivores; may contain bacteria that aid in the digestion of cellulose
Source: http://ourworld.compuserve.com/homepages/gr_frank/dig_anat.htm Cloaca:
respiration through the skin can take place in air, water, or both most important among amphibians (especially the family Plethodontidae) Gills (see Respiration in Fishes) o Cartilaginous fishes: 5 naked gill slits Anterior & posterior walls of the 1st 4 gill chambers have a gill surface (demibranch). Posterior wall of last (5th) chamber has no demibranch. Interbranchial septum lies between 2 demibranchs of a gill arch Gill rakers protrude from gill cartilage & guard entrance into gill chamber 2 demibranchs + septum & associated cartilage, blood vessels, muscles, & nerves = holobranch
o o
chamber at end of digestive tract that receives the intestine, & urinary & genital ducts, & opens to the exterior via the vent shallow or non-existent in lampreys, ray-finned fishes, & mammals (except monotremes) if no cloaca is present, the intestine opens directly to the exterior via anus
Bony fishes (teleosts):
(See 'Ventilation in Teleost Fishes')
Source: http://trc.ucdavis.edu/mjguinan/apc100/modules/Reproductive/ BIO 342 Comparative Vertebrate Anatomy Lecture Notes - Respiratory System Respiration is the process of obtaining oxygen from the external environment & eliminating CO2.
External respiration - oxygen and carbon dioxide exchanged between the external environment & the body cells Internal respiration - cells use oxygen for ATP production (& produce carbon dioxide in the process)
usually have 5 gill slits operculum projects backward over gill chambers interbranchial septa are very short or absent o Agnathans: 6 - 15 pairs of gill pouches pouches connected to pharynx by afferent branchial (or gill) ducts & to exterior by efferent branchial (or gill) ducts Larval gills: o External gills outgrowths from the external surface of 1 or more gill arches found in lungfish & amphibians o Filamentous extensions of internal gills project through gill slits occur in early stages of development of elasmobranchs o Internal gills - hidden behind larval operculum of late anuran tadpoles
Adaptations for external respiration: 1 - Primary organs in adult vertebrates are external & internal gills, swim bladders or lungs, skin, & the buccopharyngeal mucosa 2 - Less common respiratory devices include filamentous outgrowths of the posterior trunk & thigh (African hairy frog), lining of the cloaca, & lining of esophagus Respiratory organs:
Swim bladder & origin of lungs - most vertebrates develop an outpocketing of pharynx or esophagus that becomes one or a pair of sacs (swim bladders or lungs) filled with gases derived directly or indirectly from the atmosphere. Similarities between swim bladders & lungs indicate they are the same organs. Vertebrates without swim bladders or lungs include cyclostomes, cartilaginous fish, and a few teleosts (e.g., flounders and other bottom-dwellers). Swim bladders:
may be paired or unpaired (see diagram above)
Cutaneous respiration
have, during development, a pneumatic duct that usually connects to the esophagus. The duct remains open (physostomous) in bowfins and lungfish, but closes off (physoclistous) in most teleosts. serve primarily as a hydrostatic organ (regulating a fish's specific gravity) gain gas by way of a 'red body' (or red gland); gas is resorbed via the oval body on posterior part of bladder may also play important roles in: o hearing - some freshwater teleosts (e.g., catfish, goldfish, & carp) 'hear' by way of pressure waves transmitted via the swim bladder and small bones called Weberian ossicles (see diagram below) o sound production - muscles attached to the swim bladder contract to move air between 'sub-chambers' of the bladder. The resulting vibration creates sound in fish such as croakers, grunters, & midshipman fish. o respiration - the swim bladder of lungfish has number subdivisions or septa (to increase surface area) & oxygen and carbon dioxide is exchanged between the bladder & the blood
Reptilian lungs simple sacs in Sphenodon & snakes Lizards, crocodilians, & turtles - lining is septate, with lots of chambers & subchambers air exchanged via positive-pressure ventilation Avian lungs - modified from those of reptiles: air sacs (diverticula of lungs) extensively distributed throughout most of the body arrangement of air ducts in lungs ----> no passageway is a dead-end air flow through lungs (parabronchi) is unidirectional Mammalian lungs: multichambered & usually divided into lobes air flow is bidirectional:
Trachea <---> primary bronchi <---> secondary bronchi <---> tertiary bronchi <--> bronchioles <---> alveoli
Source: http://www.notcatfish.com/ichthyology/weberian_apperatus.htm Lungs & associated structures
air exchanged via negative pressure ventilation, with pressures changing due to contraction & relaxation of diaphragm & intercostal muscles
Larynx
o o o
Tetrapods besides mammals - 2 pair of cartilages: artytenoid & cricoid Mammals - paired arytenoids + cricoid + thyroid + several other small cartilages including the epiglottis (closes glottis when swallowing) Amphibians, some lizards, & most mammals - also have vocal cords stretched across the laryngeal chamber
BIO 342 Comparative Vertebrate Anatomy Lecture Notes 9 - Circulatory System Vertebrate Circulatory Systems:
Source: http://www.worldzone.net/music/singingvoice/images/glottis.gif
Trachea & syrinx o Trachea
usually about as long as a vertebrates neck (except in a few birds such as cranes) reinforced by cartilaginous rings (or c-rings) splits into 2 primary bronchi &, in birds only, forms the syrinx at that point
Lungs
Amphibian lungs 2 simple sacs internal lining may be smooth or have simple sacculations or pockets air exchanged via positive-pressure ventilation
transport gases, nutrients, waste products, hormones, heat, & various other materials consist of heart, arteries, capillaries, & veins: o Arteries carry blood away from the heart have muscular, elastic walls terminate in capillary beds o Capillaries have very thin walls (endothelium only) are the site of exchange between the blood and body cells o Veins carry blood back to the heart have less muscle in their walls than arteries but the walls are very elastic begin at the end of capillary beds o Heart a muscular pump (cardiac muscle)
contains a pacemaker to regulate rate but rate can also be influenced by the Autonomic Nervous System
1 - Heart consists of 2 atria & 2 ventricles &, except in adult birds & mammals, a sinus venosus 2 - Complete interatrial septum
Vertebrate Hearts: (see HHMI Biointeractive - click on Vertebrate circulatorium) Cartilaginous fishes
3 - Complete interventricular septum only in crocodilians, birds, & mammals; partial septum in other amniotes
single-circuit heart with 4 chambers: sinus venosus, atrium, ventricle, & conus arteriosus o the sinus venosus receives blood & is filled by suction when the ventricle contracts & enlarges the pericardial cavity o the atrium is a thin-walled muscular sac; an A-V valve regulates flow between atrium & ventricle o the ventricle has thick, muscular walls o the conus arteriosus leads into the ventral aorta (and a series of conal valves in the conus arteriosus prevent the backflow of blood)
Used by permission of John W. Kimball Arterial channels - supply most tissues with oxygenated blood (but carry deoxygenated blood to respiratory organs). In the basic pattern: 1 - the ventral aorta emerges from heart & passes forward beneath the pharynx 2 - the dorsal aorta (paired above the pharynx) passes caudally above the digestive tract 3 - six pairs of aortic arches connect the ventral aorta with the dorsal aortas
Teleosts - heart is similar to that of cartilaginous fishes, except a bulbus arteriosus (a muscular extension of the ventral aorta) is present rather than a conus arteriosus (a muscular extension of the ventricle)
Aortic arches of fishes - general pattern of development of arches in cartilaginous fishes: Used by permission of John W. Kimble 1 - Ventral aorta extends forward below pharynx & connects developing aortic arches. The first pair of arches develop first. 2 - Segments of first pair are lost & remaining sections become efferent pseudobranchial arteries 3 - Other pairs of arches (2 - 6) give rise to pre- & posttrematic arteries 4 - Arches 2 - 6 become occluded; dorsal segments = efferent branchial arteries & ventral segments = afferent branchial arteries 5 - Capillary beds develop within nine demibranchs Result: Blood entering an aortic arch from ventral aorta must pass through gill capillaries before proceeding to dorsal aorta Teleosts:
Lungfish & amphibians - modifications are correlated with the presence of lungs & enable oxygenated blood returning from the lungs to be separated from deoxygenated blood returning from elsewhere (see HHMI Biointeractive):
Partial or complete partition within atrium (complete in anurans and some urodeles) Partial interventricular septum (lungfish) or ventricular trabeculae (amphibians) to maintain separation of oxygenated & unoxygenated blood Formation of a spiral valve in the conus arteriosus of many dipnoans and amphibians. The spiral valve alternately blocks & unblocks the entrances to the left and right pulmonary arches (sending unoxygenated blood to the skin & lungs). Shortening of ventral aorta, which helps ensure that the oxygenated & unoxygenated blook kept separate in the heart moves directly into the appropriate vessels
5 = ventricle, 11 = right atrium, 12 = left atrium, 13 = conus arteriosus Amniotes:
o o
Lungfish:
the same changes convert 6 pairs of embryonic aortic arches into afferent & efferent branchial arteries arches 1 & 2 are usually lost
the pulmonary artery branches off the 6th aortic arch and supplies the swim bladder (& this is the same way that tetrapod lungs are supplied)
the other aortic trunk appears to come out of right ventricle & leads to left 4th aortic arch. So, does the left 4th arch carry oxygenated blood?
Aortic arches of tetrapods - embryos have 6 pairs of aortic arches:
o o o o o
but the are the the the the
the 1st & 2nd arches are temporary & not found in adults 3rd aortic arches & the paired dorsal aortas anterior to arch 3 called the internal carotid arteries 4th aortic arches are called the systemic arches 5th aortic arch is usually lost pulmonary arteries branch off the 6th arches & supply blood to lungs
Turtles, snakes, & lizards - the interventricular septum is incomplete where right & left systemic arches (4th) leave the ventricle & trabeculae in that region of the heart form a pocket called the cavum venosum. Oxygenated blood from the left ventricle is directed into cavum venosum, which leads to the 2 systemic arches. As a result, both the left & right systemic arches receive oxygenated blood. (see HHMI Biointeractive)
Source: http://www.uta.edu/biology/campbell/cva/3452circ.htm Further modifications of tetrapod arches: Amphibians:
Source: http://www.ulg.ac.be/physioan/chapitre/ch2s2.htm Crocodilians - ventricular septum is complete but a narrow channel called the Foramen of Panizza connects the base of the right & left systemic trunks (see HHMI Biointeractive)
o o
Urodeles - most terrestrial urodeles have 4 pairs of arches; aquatic urodeles typically have 3 pairs (III, IV, & VI) Anurans - have 4 arches early in development (larval stage); arch VI develops a pulmonary artery (to lungs) while arches III, IV, & V supply larval gills. At metamorphosis: aortic arch 5 is lost the dorsal aorta between arches 3 & 4 is lost, so blood entering arch 3 (carotid arch) goes to the head a segment (ductus arteriosus) of arch 6 is lost so blood entering this arch goes to the skin & lungs aortic arch 4 (systemic arch) on each side continue to the dorsal aorta & distributes blood to the rest of the body Oxygenated blood from the left atrium & deoxygenated blood from the right are largely kept separate in the ventricle by: Ventricular trabeculae Spiral valve in the conus arteriosus Reptiles - 3 aortic arches in adults (III, IV, & VI) Ventral aorta - no spiral valve but truncus arteriosus is split into 3 separate passages: 2 aortic trunks & a pulmonary trunk. As a result: pulmonary trunk emerges from the right ventricle & connects with 6th aortic arches (deoxygenated blood from right atrium goes to lungs) one aortic trunk comes out of left ventricle & carries oxygenated blood to the right 4th aortic arch & to carotid arches
Source: http://www.auburn.edu/academic/classes/zy/0301/Topic16/Topic16.html Role of the Foramen of Panizza in the crocodilian circulatory system:
When a crocodilian is above water and breathing air, the semilunar valve in the right aorta remains closed because of higher pressure in the left & right aorta (higher than in the right ventricle). As a result, the right aorta receives blood from the left aorta (so both aortas carry oxygenated blood) and blood from the right ventricle (low in oxygen) passes only into the pulmonary artery (and goes to the lungs).
Source: http://www.stanford.edu/~chaoyc/VL/review/problem/lf_example1.html#
When a crocodilian is under water and not breathing, right ventricular pressure increases due to pulmonary resistance (vasoconstriction of blood vessels supplying the lungs). As a result, the semilunar valve in the right aorta is now forced open so some of the blood from the right ventricle now enters the right aorta rather than the pulmonary artery. This means that, rather than going to the lungs (where there is little or no oxygen anyway because the crocodilian is under water & not breathing), some of the blood enters the systemic (body) circulation. This means that vital organs & tissues (such as skeletal muscles and the central nervous system) will get an increased blood supply and additional oxygen. This, in turn, allows a crocodilian to stay underwater longer (which is most important because many crocodilians hunt by remaining underwater and 'ambushing' prey that come for a drink or to cool off).
Oreillette droite = right atrium, Oreillette gauche = left atrium, Ventricule droit = right ventricle, Ventricule gauche = left ventricle Source: http://www.ulg.ac.be/physioan/chapitre/ch2s2.htm Secret of the crocodile heart (Franklin, C.E., and M. Axelsson. 2000. An actively controlled heart valve. Nature 406:847) By examining the heart of a crocodile, researchers have discovered how it is that an air-breathing creature can manage to cruise through the murk, for several hours without surfacing. The crocodile has a unique type of valve in its heart which actively controls blood flow between the lungs and the rest of the body. University of Queensland researcher, Craig Franklin, together with University of Goteborg colleague Michael Axelsson have been studying the heart of the estuarine crocodile, Crocodylus porosus. "These valves represent an absolute evolutionary novelty, said Dr Franklin. They are further proof of the complexity and sophistication of the 'plumbing' and general anatomy of the crocodile family," Dr Franklin said. Unlike the passive flap-like valves of other vertebrates, the crocodile valve has cog teeth made up of nodules of connective tissue. The cog teeth mesh together, diverting blood away from lungs and into their bodies. The researchers have found that these teeth are controlled by the amount of adrenalin in the bloodstream."When the crocodile is relaxed, the absence of adrenalin acts to close the cog-teeth valves," Dr Franklin said. He said this mechanism may allow the crocodiles to dive for several hours without needing to resurface to breathe. The valves are situated in the crocodile's right ventricle, which pumps blood to the pulmonary artery feeding the lungs as well as to the left aorta which supplies the body. The cog-teeth valve can divert blood going to the lungs back into the body, a phenomenon known as a shunt. "In contrast, mammalian hearts are very inflexible with the blood supply to the lungs a separate activity to that feeding the body." Abbie Thomas - ABC Science Online Birds & mammals - no mixing of oxygenated & unoxygenated blood; complete interventricular septum + division of ventral aorta into 2 trunks:
lateral abdominal hepatic portal coronary pulmonary
Source: http://www.auburn.edu/academic/classes/zy/0301/Topic16/Topic16.html The venous channels in sharks:
Cardinal streams - sinus venosus receives all blood returning to heart. Most blood enters sinus venosus via Common Cardinals. Blood from head is collected by Anterior Cardinals. Postcardinals receive renal veins & empty into Common Cardinals. Renal Portal stream - Early in development, some blood from caudal vein continues forward as Subintestinal (drains digestive system); this connection is then lost. During development, afferent renal veins (from old postcardinals) invade kidneys, & old postcardinals near top of kidneys are lost; all blood from tail must now enter kidney capillaries. Lateral Abdominal stream - LA vein starts at pelvic fin (where it receives iliac vein) & passes along lateral body wall; receives brachial vein, then turns, becomes Subclavian vein, & enters Common Cardinal vein. Hepatic Portal stream & Hepatic sinuses - Among 1st vessels to appear in vertebrate embryos are Vitelline veins (from yolk sac to heart). One Vitelline vein joins with embryonic Subintestinal vein (that drains digestive system) & becomes the Hepatic Portal System. Between liver & sinus venosus, 2 Vitelline veins are known as Hepatic sinuses.
Source: http://www.uta.edu/biology/restricted/3452circ.htm Venous channels in other fishes are much like those of sharks except:
Pulmonary trunk that takes blood to the lungs Aortic trunk that takes blood to the rest of the body
Cyclostomes have no renal portals In most bony fishes the lateral abdominals are absent & the pelvic fins are drained by postcardinals
Result of modifications: All blood returning to right side of heart goes to the lungs; blood returning from lungs to the left side of heart goes to systemic circulation.
Venous channels of tetrapods - early embryonic venous channels are very similar to those of embryonic sharks. Changes during development include:
Venous channels - In early vertebrate embryos, venous channels conform to a single basic pattern. As development proceeds, these channels are modified by deletion of some vessels & addition of others. The primary venous pathways include:
cardinals renal portal
Cardinal veins & precavae - embryonic tetrapods have posterior cardinals, anterior cardinals, & common cardinals o Urodeles - posterior cardinals persist between caudal vein & common cardinals in adults o Anurans, most reptiles, & birds - posterior cardinals are lost anterior to kidneys o Mammals - right posterior cardinal persists (azygos); part of left posterior cardinal persists (hemiazygos)
Terminology note: Common cardinals in tetrapods are called PRECAVAE; anterior cardinals are called INTERNAL JUGULAR VEINS.
efficiently as possible involves a series of vessels & openings found only in a mammalian fetus:
Some mammals (e.g., cats & humans) lose the left precava; the left brachiocephalic carries blood from left side to right precava (sometimes called SUPERIOR VENA CAVA). Postcava - Both posterior cardinals begin to develop in embryos, but only one persists & becomes the POSTCAVA. The Postcava passes directly through the liver (sort of an expressway for blood from kidneys & the posterior part of the body to the heart). The Postcava is sometimes called the INFERIOR VENA CAVA. In crocodilians, birds, & mammals, veins from hindlimbs connect directly to Postcava. Abdominal stream: o Early tetrapod embryos - paired lateral veins (like lateral abdominals of sharks) begin in caudal body wall near hind limbs, continue cranially, receive veins from forelimbs, & empty into cardinal veins or sinus venosus. As development continues: Amphibians - 2 abdominal veins fuse at midventral line & form VENTRAL ABDOMINAL VEIN. Blood in this vessel goes into liver capillaries & abdominals anterior to liver are lost (so abdominal stream no longer drains anterior limbs). Reptiles - 2 lateral abdominals do not fuse but still terminate in liver capillaries (so do not drain anterior limbs; see diagram below). Birds - retain none of their embryonic abdominal stream as adults Mammals - no abdominal stream in adults Renal Portal system: o Amphibians & some reptiles - acquires a tributary (external iliac vein; not homologous to mammalian external iliac) which carries some blood from the hind limbs to the renal portal vein. This channel provides an alternate route from the hind limbs to the heart. o Crocodilians & birds - some blood passing from hind limbs to the renal portal by-passes kidney capillaries, going straight through the kidneys to the postcava (see diagram above) o Mammals - renal portal system not present in adults Hepatic Portal system - similar in all vertebrates; drains stomach, pancreas, intestine, & spleen & terminates in capillaries of liver Pulmonary veins - carry blood from lungs to left atrium in lungfish & tetrapods
blood (with oxygen & nutrients acquired in placenta) passes into umbilical vein blood largely bypasses the liver via the ductus venosus blood returns to the heart & enters right atrium, but much of the blood then bypasses the right ventricle & enters the left atrium via the foramen ovale blood that does enter the right ventricle largely bypasses the pulmonary circulation via the ductus arteriosus
Major changes at birth: 1 - Ductus arteriosus closes 2 - Foramen ovale sealed off 3 - Blood no longer flows through umbilical vein Lymphatic system - found in all vertebrates; consists of lymph vessels, lymph nodes, &, in some species, lymph hearts
Lymph vessels o found in most soft tissues of the body & begin as blind-end lymph capillaries that collect interstitial fluid o valves present (in birds & mammals) that prevent backflow o empty into 1 or more veins (e.g., caudal, iliac, subclavian, & posterior cardinal) Lymph nodes - located along lymph vessels; contain lots of lymphocytes & macrophages (phagocytic cells) Lymph hearts - consist of pulsating smooth muscle that propels lymph fluid through lymph vessels; found in fish, amphibians, & reptiles
BIO 342 Comparative Vertebrate Anatomy Lecture Notes 10 - Urogenital System Vertebrate kidneys consist of glomeruli, tubules surrounded by peritubular capillaries, & longitudinal ducts. Variations in kidney structure among vertebrates are primarily in the number & arrangement of the glomeruli & tubules.
Circulation in a mammalian fetus & changes at birth:
In a developing fetus, blood obtains oxygen (& gives up carbon dioxide) via the placenta, not the lungs. As a result, blood flow must largely bypass the lungs so that oxygentated blood can get to other developing tissues. Getting oxygenated blood from the placenta back to the heart & out to the body as quickly and
Glomeruli are masses of capillaries that (along with Bowman's capsule) 'filter' the blood - the first step in eliminating waste products from the blood. Kidney tubules collect the glomerular filtrate & conduct it to a longitudinal duct. Tubules consist of several segments & begin as a Bowmans (or
glomerular) capsule. A glomerulus plus its surrounding Bowman's capsule is called a renal corpuscle. Longitudinal ducts = begin developing at anterior end of kidney & grow caudally until opening into the cloaca
the adult amniote kidney the number of corpuscles is large; up to about 4.5 million is some species drained by a duct called the metanephric duct or ureter
Mammalian kidneys are divided into the CORTEX(#5), MEDULLA (#6), & PELVIS(#4): 1 = Bowman's capsule, 2 = glomerulus, 3 = afferent arteriole, 4 = efferent arteriole, 5 = proximal convoluted tubule, 6 = distal convoluted tubule, 7 = collecting duct, 8 = loop of Henle, 9 = peritubular capillaries
Cortex - contains renal corpuscles & lots of capillaries Medulla - contains collecting ducts and loops of Henle; divided into pyramids (#7) & columns (#2) Pelvis - hollow; receives the urine (which exits the kidney via the ureter #3)
Archinephros - earliest vertebrate kidneys probably extended the entire length of the body cavity & had external glomeruli that drained the coelomic fluid Pronephros - 1st embryonic tubules in all vertebrates; called pronephric tubules because they are the 1st to develop & are anteriorly located
Tubules of mammalian kidney have U-shaped Loops of Henle (avian kidney = very short loops & reptilian kidney = no loops) Blood supply:
Number - never very many (e.g., 3 in frogs, 7 in human embryos, & 12 in chicken embryos) The duct that drains the pronephros is called the pronephric duct. The pronephros is temporary & function only until glomeruli & tubules further back become functional.
kidney is supplied by 2 or more renal arteries in reptiles & birds, & by a single renal artery in mammals (below). Pathway of blood in mammalian kidney: renal artery > segmental arteries > interlobar arteries > arcuate arteries > interlobular arterioles.
Urinary bladders are found in all vertebrates except agnathans, snakes, crocodilians, some lizards, & birds (except ostriches).
Mesonephros:
formed by corpuscles & tubules that develop caudal to pronephric region; form connections with existing pronephric duct (which is now called the mesonephric duct) the embryonic kidney in reptiles, birds, & mammals the functional adult kidney in fish & amphibians (& sometimes called the opisthonephros)
Fish - bladders are terminal enlargements of the mesonephric ducts called TUBAL BLADDERS Amphibians through Mammals - bladders arise as evaginations of ventral wall of the cloaca
Value of tetrapod urinary bladder:
Jawed fishes & amphibians - among males, some anterior tubules of mesonephros conduct sperm from testis to mesonephric duct. That part of the mesonephros is called the SEXUAL KIDNEY while the rest is the URINIFEROUS KIDNEY. Amniote embryos - mesonephros functions for a short time after hatching or birth &, during that time, a new kidney called the metanephros is developing
void urine when desired rather than continuously as it is formed uses of urine: o reproduction (e.g., providing males with information concerning the reproductive status of a female) o behavioral (e.g., marking territories) o moisten soil (some freshwater turtles use urine to soften the ground and make it easier to dig holes for egg-laying)
Gonads:
Metanephros:
arise as paired ridges just medial to mesonephroi
due to fusion or failure of 1 ridge to differentiate, some vertebrates (agnathans, some female lizards & crocodilians, & most female birds) have a single testis or ovary hormones cause differentiation of early gonads into either testes or ovaries
useful when fertilization is internal; introduce sperm into female reproductive tract found in some fish, some birds, reptiles, & mammals o cartilaginous fish - appendages of pelvic fins called claspers direct sperm into female reproductive tract
Ovaries:
o
Testes: In some teleosts, ovaries are hollow sacs, either because the ovary develops around coelom or the ovary becomes hollow at ovulation (eggs are discharged into cavity which is continuous with the oviduct) In other teleosts plus agnathans, the ovaries are compact & eggs are discharged into coelom Amphibians - ovaries are hollow & eggs are discharged into the coelom Reptiles, birds, & monotremes - ovaries solid but develop irregular, fluidfilled lacunae (cavities); eggs discharged into coelom Mammals - ovaries compact; no large chambers or lacunae
snakes & lizards - have pair of HEMIPENES (pocketlike diverticula of wall of cloaca) turtles, crocodilians, a few birds, & mammals - exhibit an unpaired erectile penis penis - usually a thickening of floor of cloaca consisting of spongy erectile tissue (corpus spongiosum) with grooves to direct sperm & ending in a glans penis (sensory endings that reflexly stimulate ejaculation) mammals (except monotremes) - penis extends beyond body. The embryonic corpus spongiosum becomes a tube with urethra inside & 2 additional erectile masses develop (corpus cavernosa).
Female genital ducts: usually smaller than ovaries because sperm, although numerous, are much smaller than eggs (especially eggs with yolk) In mammals, testes are larger than ovaries
Translocation of testes in mammals:
typically consists of a pair of gonoducts (or oviducts) that extend from ostia to the cloaca different segments of ducts perform special functions when internal, fertilization occurs near beginning of ducts
testes descend permanently into scrotal sacs in many mammals some mammals - testes lowered into scrotal sacs & retracted at will inguinal canal - passage between abdominal cavity & scrotum scrotal sacs - do not develop in some mammals; testes remain in abdomen
Source: http://gwis2.circ.gwu.edu/~atkins/newwebpages/Reproductive/breed.html
Male genital ducts:
Some fishes (e.g., gar & sturgeon) & amphibians - mesonephric duct transmits sperm & urine Some amphibians - mesonephric duct transports only sperm; new accessory urinary duct drains the kidney Sharks - mesonephric duct is used primarily for sperm transport; accessory urinary duct develops Teleosts - mesonephric duct drains kidney; separate sperm duct develops Amniotes - embryonic mesonephric ducts transport sperm in adults
anatomy in various vertebrate groups: o cartilaginous fish - 2 ostia fuse to form single ostium (or osteum); shell gland secretes albumen & a shell; uterus holds eggs until laying o teleosts - ducts are continuous with cavity of the ovary o lungfish & amphibians - oviducts long & convoluted; lining secretes jelly-like material around each egg o crocodilians, some lizards, & nearly all birds (diagram below) - 1 coiled oviduct lined with glands that add albumen, shells, &, sometimes, pigment
o
Source: http://gwis2.circ.gwu.edu/~atkins/newwebpages/Reproductive/breed.html Intromittent organs:
monotremes - tract is reptilian; caudal end secretes a shell before egg passes into the cloaca placental mammals - embryonic ducts give rise to oviducts, uteri, & vaginas. Adult tract is paired anteriorly & unpaired posteriorly (typically terminating as an unpaired vagina).
oviducts (fallopian tubes) are relatively short, small in diameter, convoluted, & lined with cilia; begin at ostium bordered with fimbria uterus: Marsupials - no fusion of embryonic ducts so there are 2 tracts (DUPLEX UTERUS)
Source: http://www.david.curtis.care4free.net/neurons.htm Neurons (or nerve cells):
Other placental mammals - varying degrees of fusion: bipartite uterus - 2 uterine horns, a uterine body (with 2 lumens), & a single vagina bicornuate uterus - 2 uterine horns, a uterine body (with a single lumen), & a single vagina simplex uterus - no uterine horns & oviducts open directly into body of uterus
respond to stimuli & conduct impulses 3 types - all with cell body & processes (axons & dendrites): o multipolar o bipolar o unipolar Bipolar neuron Unipolar neuron
Multipolar neuron Nerves = bundles of nerve cell processes; may be sensory, motor, or mixed Spinal cord:
Vagina - fused terminal portion of oviducts that opens either into urogenital sinus or to the exterior; receives male intromittent organ BIO 342 Comparative Vertebrate Anatomy Lecture Notes 11 - Nervous System The Vertebrate Nervous System: 1 - receives stimuli from receptors & transmits information to effectors that respond to stimulation 2 - regulates behavior by integrating incoming sensory information with stored information (the results of past experience) & translating that into action by way of effectors 3 - includes billions of nerve cells (or neurons), each of which establishes thousands of contacts with other nerve cells 4 - also includes neuroglia cells that support, nourish, & insulate neurons Subdivisions of the Vertebrate Nervous System: 1 - Central Nervous System - including the brain & spinal cord 2 - Peripheral Nervous System - including cranial nerves, spinal nerves, & all branches of cranial & spinal nerves
located in vertebral canal anatomical beginning is the foramen magnum of the skull length varies among vertebrates: o in vertebrates with abundant tail musculature, the spinal cord extends to the caudal end of the vertebral column o in vertebrates without tails or without much tail musculature, the spinal cord extends to about the lumbar region of the vertebral column a cross-section of the spinal cord reveals gray matter & white matter. The gray matter consists of nerve cell bodies, while the white matter consists of nerve cell processes (axons). These processes make up ascending (sensory) and descending (motor) fiber tracts.
Used with permission of G. Mandl Spinal nerves:
arise from spinal cord by dorsal & ventral roots. The dorsal root exhibits a ganglion & is sensory, while the ventral root has no ganglion & is motor. early vertebrates: o dorsal & ventral roots did not unite o dorsal roots were mixed (contained both sensory & motor fibers) o no dorsal root ganglion Rami - 2 branches of each spinal nerve: o dorsal ramus - supplies epaxial muscles & skin of the dorsal part of the body o ventral ramus - supplies hypaxial muscles & skin of the side & ventral part of the body
Functional types of neurons in spinal nerves (& other nerves): o somatic afferent - sensory from general cutaneous receptors (in the skin) & proprioceptors (in skeletal muscles, tendons, & joints) o somatic efferent - motor to skeletal muscles o visceral afferent - sensory from receptors in the viscera (smooth muscle, cardiac muscle, & glands) o visceral efferent - motor to smooth muscle, cardiac muscle, & glands
Cerebellum - modifies & monitors motor output: o important in maintaining equilibrium o coordinates & refines motor action
Mesencephalon - consists of the tectum which includes the optic lobes & auditory lobes:
Brain:
optic lobes - receive fibers from retina; vary in size with relative importance of vision auditory lobes - receive fibers from inner ear
the anterior end of the embryonic central nervous system exhibits 3 primary sections: o prosencephalon (forebrain) - subsequently divides into the telencephalon (cerebrum) & diencephalon (epithalamus, thalamus, & hypothalamus) o mesencephalon (midbrain) - develops without further subdivision & forms the tectum o rhombencephalon (hindbrain) - subdivides into the metencephalon (pons & cerebellum) and myelencephalon (medulla oblongata)
Diencephalon - consists of the epithalamus, hypothalamus, & thalamus:
epithalamus - includes pineal gland (epiphysis) that affects skin pigmentation (by acting on melanocytes) in lower vertebrates & plays a role in regulating biological rhythms in higher vertebrates hypothalamus - regulates body temperature, water balance, appetite, blood pressure, sexual behavior, & some aspects of emotional behavior thalamus - major coordinating, or relay, center for sensory impulses from all parts of the body
Source: http://brainmuseum.org/development/index.html
Source: http://www.colorado.edu/epob/epob3730rlynch/image/figure5-4.jpg Telencephalon - consists of the cerebrum which, in turn, consists of 2 cerebral hemispheres
Phylogenetic trend in vertebrate brains is for enlargement of forebrain: o increasingly complex behaviors & muscle control: coordination of limb movements more complicated (e.g., bipedal dinosaurs & birds) increased input of sensory information & increased output of motor responses
Source: http://www.colorado.edu/epob/epob3730rlynch/image/figure5-1.jpg Myelencephalon - consists of the medulla oblongata & its major functions include:
cerebrum has 2 regions: a dorsal PALLIUM (with medial, dorsal, & lateral divisions) & a ventral SUBPALLIUM (consisting of a striatum & a septum) all vertebrates have a cerebrum based on the same basic plan; major phylogenetic changes are due to loss, fusion, or enlargement of the various regions. o medial pallium receives olfactory information o dorsal & lateral pallia receive other sensory input (including visual & auditory information relayed from the thalamus) agnathans, fish, & amphibians - pallia are similar
Source: http://www.auburn.edu/academic/classes/zy/0301/Topic19/Topic19.html
origin of cranial nerves (VII - X or VII - XII) pathway for ascending & descending fiber tracts contains centers important in regulating respiration, heartbeat, & intestinal motility
Metencephalon - consists of the pons & cerebellum:
reptiles - pallium has 3 main divisions (medial, dorsal, & lateral) but also has a large DORSAL VENTRICULAR RIDGE (DVR), derived from lateral pallium; DVR may be higher association area birds - DVR expands further; dorsal part increases in size & is called the WULST; as in reptiles, the DVR appears to serve as a higher association area
Pons - pathway for ascending & descending fiber tracts & origin of cranial nerves V, VI, & VII
Source: http://www.pigeon.psy.tufts.edu/avc/husband/avc2amnt.htm
mammals - do not have enlarged DVR but DORSAL PALLIUM is enlarged & is called the CEREBRAL CORTEX; cortex receives & analyzes sensory information & initiates motor activity subpallium: o septum - important part of the limbic system (regulates emotions & plays vital role in short-term memory) o striatum - also called basal ganglia; present in all vertebrates & controls sequence of actions in complex movements
special sensory organs - confined to the head (amniotes & terrestrial amphibians)
Special Somatic Receptors
Cranial nerves - agnathans, most fish, & living amphibians have 10 cranial nerves; crossopterygians & amniotes have 12:
Neuromast organs ('groove organ' below in Figure 10-4) o receptors in skin of fishes & aquatic amphibians that detect water currents & hear sounds o occur singly, in groups, or in a linear series (e.g., lateral lines) o may also be modified to detect electricity (ampullae of Lorenzini)
Olfactory nerve (I) - sensory nerve; sense of smell Optic nerve (II) - sensory nerve; sense of vision Oculomotor nerve (III) - motor nerve to extrinsic eye muscles Trochlear nerve (IV) - motor to extrinsic eye muscles Trigeminal (V) - mixed nerve; sensory from skin of head & mouth (including teeth) & motor to muscles of 1st pharyngeal arch (muscles of jaw) Abducens (VI) - motor to extrinsic eyeball muscles Facial (VII) - mixed nerve; sensory from lateral line of head, ampullae of Lorenzini, & taste buds; motor to muscles of hyoid arch Auditory (VIII) - sensory from inner ear (balance & hearing) Glossopharyngeal (IX) - mixed nerve; sensory from taste buds & lateral line; motor to muscles of 3rd arch Vagus (X) - mixed nerve; sensory from & motor to heart, anterior digestive system, mouth, gill pouches 2 - 5, & lateral line Accessory nerve (XI) - motor to derivatives of cucullaris muscle (cleidomastoid, sternomastoid, & trapezius) Hypoglossal nerve (XII) - motor to hyoid & tongue muscles
Shark lateral line system
The ampullae of Lorenzini are small vesicles that form part of an extensive subcutaneous sensory network system. These vesicles are found around the head of the shark. They detect weak magnetic fields produced by other fish at short ranges. This enables a shark to locate prey buried in the sand or to orient to nearby movement. Each ampulla is a bundle of sensory cells innervated by several nerve fibers. These fibers are enclosed in a jelly-filled tubule that has a direct opening to the surface through a pore. These pores on the head of the shark are visible to the naked eye, and appear as dark spots.
Neuromast organs have 2 types of cells: o hair cells (receptor cells) - each hair cell has several short cilia & kinocilia that project into fluid or a cupula (displacement of cupula & cilia generates nervous impulses) o supporting cells Membranous labyrinth o exhibited by all vertebrates o fluid-filled & embedded in skull lateral to hindbrain o Labyrinth usually consists of 3 semicircular canals, a utriculus, & a sacculus
Possibly useful mnemonics to aid in memorization of cranial nerves: On Old Olympus Towering Top A Finn And German Viewed A Hop or Oh, Once One Takes The Anatomy Final, A Good Vacation Appears Heavenly.
Sensory Organs Sensory receptors:
Semicircular canals:
monitor the external & internal environment by responding to selected stimuli, then translating those stimuli into nerve impulses Types of sensory organs: o somatic sensory organs - provide information about the external environment o visceral sensory organs - provide information about the organism's internal environment o general sensory organs - widely distributed over the surface & interior of the body
Hagfish - have only one (posterior) Lamprey - have 2 (anterior & posterior) Other vertebrates - have 3 (anterior, posterior, & horizontal)
Functions of the labyrinth: 1 - Equilibrium
Dynamic equilibrium - when head moves, inertia causes a slight relative movement of fluid in at least one semicircular canal ---> deflects cupula (in ampulla) ---> nervous impulses Static equilibrium - maculae (in sacculus & utriculus) tilt when head moves ---> nervous impulses
found in pythons (Family Boidae); nerve endings lie at the bottom of several recessed labial pits permit detection of a mouse about 15 cm away
2 - Loreal pits
2 - Hearing - function of ORGAN OF CORTI located in lagena (enlargement of sacculus); lagena tends to be longer in terrestrial vertebrates &, in most mammals, its coiled into the cochlea. The organ of Corti contains a specialized strip of neuromasts connected to the nervous system via the auditory nerve.
also called facial pits; can detect temperature changes of as little as 0.001 degree C & so can detect prey several feet away present in snakes in the family Crotalidae (North American rattlesnakes, copperheads, & water moccasins), also called the pit vipers
1-Inner hair cell, 2-Outer hair cells, 3-Tunnel of Corti, 4-Basilar membrane, 5Reticular lamina, 6-Tectorial membrane, 7-Deiters' cells, 8-Space of Nuel, 9-Hensen's cells, & 10Inner spiral sulcus [Drawing by Stephan Blatrix, from "Promenade around the cochlea" by R Pujol, S. Blatrix, T. Pujol and V. Reclar-Enjalbert, CRIC, University Montpellier 1 - INSERM. URL: http://www.iurc.montp.inserm.fr/cric/audition/english/corti/fcorti.htm] Hearing:
Light receptors (or photoreceptors) - vertebrates can perceive only a narrow band of electromagnetic radiation between about 350 & 760 nm; 2 types include the epiphysis (already described) & the eye Structure of a vertebrate eye: Accommodation is the process of focusing light on the retina & this can occur in several different ways:
Outer ear of tetrapods: o Amphibians & most reptiles - eardrum (tympanic membrane) is on surface of the head o Crocodilians, birds, & mammals - eardrum is deeper in the skull at the end of an air-filled passageway called the outer ear canal (or external auditory meatus) o Mammals - pinna collects & directs sound waves Middle ear of tetrapods - cavity plus ossicle(s): o Amphibians, reptiles, & birds - single middle ear ossicle (columella or stapes) o Mammals - 3 middle ear ossicles (malleus, incus, & stapes) Inner ear = labyrinth, including lagena (or cochlea) How do pressure or sound waves become sound?
Lamprey - contraction of corneal muscle pulls cornea against the lens & moves the lens Teleosts (bony fish) - retractor muscle attached to lens (rectractor lentis muscle) moves lens posteriorly Amphibians & cartilaginous fish - protractor muscle attached to lens pulls the lens forward for near vision Snakes - increased pressure in the vitreous humor generated by muscles near the iris pushes the lens forward Most reptiles, birds, & mammals - curvature of lens is altered by ciliary (annular) muscles
Special visceral receptors - olfactory (smell) & gustatory (taste):
(Also: check out http://www.iurc.montp.inserm.fr/cric/audition/english/ear/fear.htm & http://www.blackwellscience.com/matthews/ear.html) Pit receptors of reptiles = infrared receptors: 1 - Labial pits
Olfaction - involves receptors located in nasal passages; olfactory epithelium contains basal cells (replacement cells), supporting cells (secrete mucus), & olfactory receptor cells vomeronasal organs - only in tetrapods but absent in most turtles, crocodiles, birds, some bats, primates, & aquatic mammals: o amphibians - recessed area off the main nasal cavity o reptiles - separate pit to which tongue & oral membranes deliver chemicals o mammals: isolated area of olfactory membrane within nasal cavity that is connected to mouth via a nasopalatine duct
well-developed in monotremes, marsupials, insectivores, & many carnivores function of vomeronasal organs - may be especially important in detecting conspecific odors, but also useful in prey detection
Gustation (taste) - taste buds, like olfactory receptors, detect chemical stimuli
Taste buds consist of supportive cells & taste cells distribution: Fish - widely distributed in roof, walls, & floor of pharynx; bottom feeders & scavengers (catfish & carp) have taste buds distributed over entire surface of head & body, especially on the barbels (whiskers) Tetrapods - taste buds restricted to tongue, posterior palate, & oral pharynx
cutaneous receptors (for touch, pressure, pain, & temperature) o naked endings - in skin of all vertebrates; stimulated by contact o encapsulated endings - present in tetrapods; nerve endings wrapped in a connective tissue capsule Herbst corpuscles - on beak, tongue, & palate of water birds end bulbs & Ruffini corpuscles - thermal receptors in mammals Pacinian corpuscles - touch & pressure receptors proprioceptors - located in skeletal muscles, joints, and tendons & provide information about body position
General Visceral Receptors:
General Somatic Receptors - come in two categories: cutaneous receptors & proprioceptors
mostly naked endings in mucosa of the tubes, vessels, & organs of the body, in cardiac muscle,& in smooth muscle; chiefly stretch & chemoreceptors some functions of general visceral receptors: o monitor oxygen & carbon dioxide content of blood o monitor blood pressure (baroreceptors in Figure 1 below) o monitor concentration of solutes in blood similar among all vertebrates
Vous aimerez peut-être aussi
- Comparative Vertebrate Anatomy ReviewerDocument3 pagesComparative Vertebrate Anatomy ReviewerJohn Rudolf CatalanPas encore d'évaluation
- Comparative Anatomy of Vertebrate SkeletonDocument53 pagesComparative Anatomy of Vertebrate SkeletonYlane Lee100% (1)
- Skeletal System Zoo 102Document14 pagesSkeletal System Zoo 102Rosselle NoyoalPas encore d'évaluation
- Circulation and Respiration Lungfishes (Dipnoi) : W B .) and DepartmentDocument20 pagesCirculation and Respiration Lungfishes (Dipnoi) : W B .) and DepartmentPebrianti putriPas encore d'évaluation
- Human AnatomyDocument7 pagesHuman AnatomyMitko0% (1)
- Video Worksheet Great TransDocument4 pagesVideo Worksheet Great Transb-ahmasaniPas encore d'évaluation
- Comparative Anatomy LecDocument2 pagesComparative Anatomy LecKerbie JaydPas encore d'évaluation
- Anatomy - Wikipedia, The Free EncyclopediaDocument17 pagesAnatomy - Wikipedia, The Free EncyclopediaalsdjflakjdfPas encore d'évaluation
- Anatomy and Physiology of Cerebrospinal Fluid 2011Document8 pagesAnatomy and Physiology of Cerebrospinal Fluid 2011Camilo Benavides BurbanoPas encore d'évaluation
- Cat Skeletal SystemDocument26 pagesCat Skeletal SystemMielah RuthPas encore d'évaluation
- Comparative Vertebrate AnatomyDocument9 pagesComparative Vertebrate AnatomyJayson Tafalla LopezPas encore d'évaluation
- Chapter 5 Comparative Vertebrate AnatomyDocument11 pagesChapter 5 Comparative Vertebrate AnatomyNathaniel Arthur EstavilloPas encore d'évaluation
- Biology 453 - Comparative Vert. Anatomy WEEK 1, LAB 1: Anatomical Terms & Introduction To Chordates AssignmentsDocument10 pagesBiology 453 - Comparative Vert. Anatomy WEEK 1, LAB 1: Anatomical Terms & Introduction To Chordates AssignmentsKerbie Jayd0% (1)
- MSC ZoologyDocument26 pagesMSC ZoologyRaji Deol BandeshaPas encore d'évaluation
- Skeletal SystemDocument47 pagesSkeletal Systemapi-3845840100% (1)
- A Model For The Evolution of Decussation PDFDocument5 pagesA Model For The Evolution of Decussation PDFGustavo CabanasPas encore d'évaluation
- 1008458-Chapter 7 - Ecosystem DynamicDocument12 pages1008458-Chapter 7 - Ecosystem DynamicrhimalinyPas encore d'évaluation
- Animals - Adaptations Endoskeleton, Echinoderms, Invert Chordates, VertebratesDocument9 pagesAnimals - Adaptations Endoskeleton, Echinoderms, Invert Chordates, VertebratessmedificationPas encore d'évaluation
- A System Based' Approach To Risk Assessment of The Cervical Spine Prior ToDocument9 pagesA System Based' Approach To Risk Assessment of The Cervical Spine Prior ToRudolfGerPas encore d'évaluation
- Comparative Anatomy of Vertebrates Presentation Lect 1Document20 pagesComparative Anatomy of Vertebrates Presentation Lect 1Ian Kenneth Cabrera100% (10)
- BIO 342 Comparative Vertebrate Anatomy Lecture Notes 2 - Vertebrate Skeletal Systems & SkullDocument76 pagesBIO 342 Comparative Vertebrate Anatomy Lecture Notes 2 - Vertebrate Skeletal Systems & SkullSteph VeePas encore d'évaluation
- Compa SkeletalDocument37 pagesCompa SkeletalJamie LiwanagPas encore d'évaluation
- Skeletal Urogenital Lecture ComparativeDocument136 pagesSkeletal Urogenital Lecture ComparativeLei Jenevive UmbayPas encore d'évaluation
- 08 Vertebra, Ribs, and SternaDocument3 pages08 Vertebra, Ribs, and SternaKaye Coleen Cantara Nuera100% (1)
- Skeletal and Muscular Systems of VertebratesDocument9 pagesSkeletal and Muscular Systems of VertebratesJacob NoraPas encore d'évaluation
- Chapter 1: Introduction: Phylum Chordata: The Big FourDocument6 pagesChapter 1: Introduction: Phylum Chordata: The Big FourKariza AbuPas encore d'évaluation
- 2.1 (A) - Support & Locomotion in Humans & AnimalsDocument61 pages2.1 (A) - Support & Locomotion in Humans & Animalsliming8112Pas encore d'évaluation
- Exercise 4: Skeletal SystemDocument8 pagesExercise 4: Skeletal SystemstevemacabsPas encore d'évaluation
- Sekolah Berasrama Penuh Integrasi Gopeng Prepared By:Muhd Fazli Bin DollahDocument60 pagesSekolah Berasrama Penuh Integrasi Gopeng Prepared By:Muhd Fazli Bin DollahrameshkasinathanPas encore d'évaluation
- Chapter 8Document4 pagesChapter 8Mariecris SeñoPas encore d'évaluation
- Vertebral Column Structure and FunctionDocument5 pagesVertebral Column Structure and FunctionBearer2008Pas encore d'évaluation
- 2 - Comparative Vertebrate AnatomyDocument35 pages2 - Comparative Vertebrate AnatomyGlen MangaliPas encore d'évaluation
- Skeletal System: The Skull - Anatomy and Functions in 40 CharactersDocument24 pagesSkeletal System: The Skull - Anatomy and Functions in 40 CharactersJessa BellePas encore d'évaluation
- Comparative Anatomy: (Chordates) Skeletal SystemDocument66 pagesComparative Anatomy: (Chordates) Skeletal SystemPAU FRANCISCOPas encore d'évaluation
- SkullDocument11 pagesSkullGeorgePas encore d'évaluation
- 2.1 (A) - Support & Locomotion in Humans & AnimalsDocument59 pages2.1 (A) - Support & Locomotion in Humans & AnimalsFauziah Abu Bakar100% (1)
- Sekolah Menengah Kebangsaan Permas Jaya: Prepared By:-Azam SenseiDocument59 pagesSekolah Menengah Kebangsaan Permas Jaya: Prepared By:-Azam SenseiazamsenseiPas encore d'évaluation
- Chordata Introduction: Phylum OverviewDocument2 pagesChordata Introduction: Phylum OverviewTrisha RosalesPas encore d'évaluation
- Mammalian Skeleton and Skull AnatomyDocument6 pagesMammalian Skeleton and Skull Anatomyajmal khanPas encore d'évaluation
- Skeletal SystemDocument52 pagesSkeletal SystemHarish musictryPas encore d'évaluation
- 2 Skeletal PDFDocument7 pages2 Skeletal PDFvada_soPas encore d'évaluation
- Skeleton Ikan IIDocument15 pagesSkeleton Ikan IIYvonee XaviersPas encore d'évaluation
- Med 8Document2 pagesMed 8Lef RJPas encore d'évaluation
- Chapter 1-Musculoskeletal SystemDocument113 pagesChapter 1-Musculoskeletal SystemWang Ming YaPas encore d'évaluation
- Skeletal SystemDocument23 pagesSkeletal SystemrosabelalebsPas encore d'évaluation
- Zoology MidtermsDocument14 pagesZoology MidtermsJULIANNE ANACTAPas encore d'évaluation
- The Skeletal SystemDocument41 pagesThe Skeletal SystemLouella ArtatesPas encore d'évaluation
- Skeletal System Animal ScienceDocument16 pagesSkeletal System Animal Sciencealexenneth canilaPas encore d'évaluation
- Session 14 Skeletal SystemDocument17 pagesSession 14 Skeletal SystemScience Of LifePas encore d'évaluation
- 7 Unikonta - Deuterostomes & ChordatesDocument5 pages7 Unikonta - Deuterostomes & ChordateskahiauPas encore d'évaluation
- Module 1 Chapters 1 and 2Document125 pagesModule 1 Chapters 1 and 2Liana Marie Castro DavidPas encore d'évaluation
- Compare Vertebrate Skeletal SystemsDocument12 pagesCompare Vertebrate Skeletal SystemsChucksters79Pas encore d'évaluation
- Locomotiom & The Human Skeletal SystemDocument17 pagesLocomotiom & The Human Skeletal SystemNathanael MfumbaikaPas encore d'évaluation
- 09+AppendicularDocument45 pages09+AppendicularnayoonPas encore d'évaluation
- 2.1 (A) - Support & Locomotion in Humans & AnimalsDocument59 pages2.1 (A) - Support & Locomotion in Humans & AnimalsilyPas encore d'évaluation
- Lecture Note - Skull and Visceral SkeletonDocument4 pagesLecture Note - Skull and Visceral SkeletonLopez Manilyn CPas encore d'évaluation
- Comparative Vertebrate Anatomy Lecture 1Document76 pagesComparative Vertebrate Anatomy Lecture 1Mielah Ruth100% (1)
- 2.1 (A) - Support & Locomotion in Humans & AnimalsDocument59 pages2.1 (A) - Support & Locomotion in Humans & Animalsliefeng81100% (8)
- Frog SkeletonDocument2 pagesFrog SkeletonJermaine Joyce Arvesu-Lugada0% (2)
- Lab Rep 4 FinalDocument21 pagesLab Rep 4 FinalBella LopezPas encore d'évaluation
- 3 H6P 3 F6P 2Fbp Hps Phi PFKDocument1 page3 H6P 3 F6P 2Fbp Hps Phi PFKHazel Grace BellenPas encore d'évaluation
- RA 6713 - Code of Conduct and Ethical Standard For Public Officials and EmployeesDocument12 pagesRA 6713 - Code of Conduct and Ethical Standard For Public Officials and EmployeesCrislene Cruz83% (12)
- Philippines Civil Service Professional Reviewer - Part 1Document24 pagesPhilippines Civil Service Professional Reviewer - Part 1Michael L. Dantis50% (2)
- Photo MorphogenesisDocument18 pagesPhoto MorphogenesisHazel Grace BellenPas encore d'évaluation
- Hormones 2Document17 pagesHormones 2Hazel Grace BellenPas encore d'évaluation
- Animal RespirationDocument56 pagesAnimal RespirationHazel Grace BellenPas encore d'évaluation
- Biotech Chapter 5Document21 pagesBiotech Chapter 5Hazel Grace BellenPas encore d'évaluation
- GradesDocument3 pagesGradesHazel Grace BellenPas encore d'évaluation
- Patterns of Plant DevelopmentDocument11 pagesPatterns of Plant DevelopmentHazel Grace BellenPas encore d'évaluation
- Biochemistry of Hormones 1Document21 pagesBiochemistry of Hormones 1Hazel Grace BellenPas encore d'évaluation
- Biochemistry of Hormones 2Document42 pagesBiochemistry of Hormones 2Hazel Grace BellenPas encore d'évaluation
- Cat BonesDocument12 pagesCat BonesHazel Grace BellenPas encore d'évaluation
- Clone WarsDocument3 pagesClone WarsHazel Grace BellenPas encore d'évaluation
- StatisticsDocument8 pagesStatisticsHazel Grace BellenPas encore d'évaluation
- Journal ReadingDocument4 pagesJournal ReadingHazel Grace BellenPas encore d'évaluation
- Presentation and Collection of DataDocument48 pagesPresentation and Collection of DataHazel Grace BellenPas encore d'évaluation
- Guide To Writing A Basic EssayDocument7 pagesGuide To Writing A Basic EssayDarkPrincessPas encore d'évaluation
- Rizal IssuesDocument4 pagesRizal IssuesHazel Grace BellenPas encore d'évaluation
- Cat Dissection Guide NetDocument46 pagesCat Dissection Guide NetHazel Grace Bellen100% (3)
- Cranial Base AnatomyDocument14 pagesCranial Base AnatomyBahadur Baloch100% (1)
- Pattern of Preservation of Human - Classificacao Tafonomia - Bello and AndrewsDocument13 pagesPattern of Preservation of Human - Classificacao Tafonomia - Bello and Andrewsyuri silvaPas encore d'évaluation
- Exercise 1 Frog SkeletonDocument16 pagesExercise 1 Frog SkeletonMargarette IsaacPas encore d'évaluation
- Skull Anatomy of the Magellanic PenguinDocument15 pagesSkull Anatomy of the Magellanic PenguinmedePas encore d'évaluation
- SkullDocument98 pagesSkullAl-ajim HadjiliPas encore d'évaluation
- Veroni Et Al-2010-American Journal of Physical AnthropologyDocument5 pagesVeroni Et Al-2010-American Journal of Physical AnthropologyT.Pas encore d'évaluation
- Cunninghams Manual of Practical Anatomy v3 1000902964Document624 pagesCunninghams Manual of Practical Anatomy v3 1000902964adiseifPas encore d'évaluation
- The Essential Identity of The Klippel-Feil Syndrome and IniencephalyDocument16 pagesThe Essential Identity of The Klippel-Feil Syndrome and IniencephalyIsolda Alanna RlPas encore d'évaluation
- Chapter 1 - Adjectival Forms Chapter 1 - Adjectival FormsDocument8 pagesChapter 1 - Adjectival Forms Chapter 1 - Adjectival FormsQum ArPas encore d'évaluation
- Nonsyndromic CraniosynostosisDocument10 pagesNonsyndromic CraniosynostosisOWintero OWarmoPas encore d'évaluation
- Posterior Cranial Fossa Anesthetic ManagementDocument48 pagesPosterior Cranial Fossa Anesthetic ManagementDivya Rekha KolliPas encore d'évaluation
- Week 03a Insect HeadDocument5 pagesWeek 03a Insect HeadVanshika ChaudharyPas encore d'évaluation
- Thomas Rakosi CefalometrieDocument224 pagesThomas Rakosi CefalometrieMario Mutuleanu100% (7)
- Skull BonesDocument25 pagesSkull Bonesjohn mwambuPas encore d'évaluation
- Arab Board Final 2018 - 2Document12 pagesArab Board Final 2018 - 2lonsilord17Pas encore d'évaluation
- The Skeleton: Dr. Ali EbneshahidiDocument64 pagesThe Skeleton: Dr. Ali EbneshahidiMarcus Randielle FloresPas encore d'évaluation
- AOI APState August10Document141 pagesAOI APState August10Anoop BabuPas encore d'évaluation
- The Skull and Vicsceral Skeleton PDFDocument13 pagesThe Skull and Vicsceral Skeleton PDFAmberValentinePas encore d'évaluation
- Skull Bones GuideDocument17 pagesSkull Bones Guideمدونة الأحترافPas encore d'évaluation
- The Aqueduct of Sylvius A Sonographic Landmark ForDocument7 pagesThe Aqueduct of Sylvius A Sonographic Landmark Forcarlos cobeloPas encore d'évaluation
- Bio 201 - Bone Practical Part 1 (Axial Skeleton)Document5 pagesBio 201 - Bone Practical Part 1 (Axial Skeleton)Gretchen100% (1)
- Skull Bones and LandmarksDocument8 pagesSkull Bones and LandmarksbangityPas encore d'évaluation
- CRANIECTOMY OR RotationDocument41 pagesCRANIECTOMY OR RotationFretzgine Lou ManuelPas encore d'évaluation
- A Standardized Nomenclature For Craniofacial and Facial AnthropometryDocument17 pagesA Standardized Nomenclature For Craniofacial and Facial AnthropometryxavicojmPas encore d'évaluation
- HUMAN ANATOMY: SKELETAL SYSTEMDocument15 pagesHUMAN ANATOMY: SKELETAL SYSTEMKert trocioPas encore d'évaluation
- Skull Reviewer 2Document6 pagesSkull Reviewer 2Christian Bermas DioPas encore d'évaluation
- Anatomy of the SkullDocument29 pagesAnatomy of the Skullaya abdullahPas encore d'évaluation
- Head Massage TherapyDocument169 pagesHead Massage TherapyAnonymous vaQaeG15f100% (1)
- Thescelosaurus assiniboiensis sp. nov. Brown et al., 2011. a New Basal Ornithopod Dinosaur (Frenchman Formation, Saskatchewan Canada) Canada), and implications for late Maastrichtian ornithischian diversity in North AmericaDocument42 pagesThescelosaurus assiniboiensis sp. nov. Brown et al., 2011. a New Basal Ornithopod Dinosaur (Frenchman Formation, Saskatchewan Canada) Canada), and implications for late Maastrichtian ornithischian diversity in North AmericanoaraptorPas encore d'évaluation
- OMM Fellows ReviewDocument178 pagesOMM Fellows ReviewTony Ziherl100% (1)